1-aminocyclopro-pane-1-carboxylate synthase protein turnover in Arabidopsis
1
14
... 乙烯的信号转导及功能受到泛素-蛋白酶体系统的调节[62].ACS作为乙烯生物合成的关键限速酶,其蛋白丰度受到ETO1(ethylene overproducing 1)及其同源蛋白EOL1(ETO1-like)和EOL2的调控[63].ETO1/EOLs是CULLIN3 E3泛素连接酶的组分,可以特异性识别2型ACS并通过26S蛋白酶体进行快速降解[63].14-3-3蛋白是真核生物中高度保守的二聚体蛋白,通过磷酸化介导与靶蛋白相互作用参与多种生物过程[64].在碳饥饿条件下,拟南芥出现叶片黄化衰老的表型,同时,碳饥饿会激活油菜素甾醇(brassinosteroid, BR)信号通路,由BR诱导的磷酸化的SINAT2(seven-in-absentia of A. thaliana 2)和ETO1/EOL2将14-3-3蛋白招募到复合物中,激活SINAT2与EOL2的降解,增加ACS5蛋白的丰度和自噬活性,最终促进乙烯生物合成以响应碳饥饿[65].24-表油菜素内酯(24-epibrassinolide, EBR)是BR家族中的一种.有报道发现,EBR可增加富铁幼苗根系高铁还原酶FRO的活性,却降低了缺铁幼苗中FRO的活性;进一步研究发现,EBR抑制缺铁诱导的乙烯产生,但促进富铁幼苗中乙烯的产生,说明BR可能通过与乙烯交互负调控FRO活性,从而参与植物对缺铁的响应[66]. ...
Reciprocal antagonistic regulation of E3 ligases controls ACC synthase stability and responses to stress
1
2021
... 乙烯的信号转导及功能受到泛素-蛋白酶体系统的调节[62].ACS作为乙烯生物合成的关键限速酶,其蛋白丰度受到ETO1(ethylene overproducing 1)及其同源蛋白EOL1(ETO1-like)和EOL2的调控[63].ETO1/EOLs是CULLIN3 E3泛素连接酶的组分,可以特异性识别2型ACS并通过26S蛋白酶体进行快速降解[63].14-3-3蛋白是真核生物中高度保守的二聚体蛋白,通过磷酸化介导与靶蛋白相互作用参与多种生物过程[64].在碳饥饿条件下,拟南芥出现叶片黄化衰老的表型,同时,碳饥饿会激活油菜素甾醇(brassinosteroid, BR)信号通路,由BR诱导的磷酸化的SINAT2(seven-in-absentia of A. thaliana 2)和ETO1/EOL2将14-3-3蛋白招募到复合物中,激活SINAT2与EOL2的降解,增加ACS5蛋白的丰度和自噬活性,最终促进乙烯生物合成以响应碳饥饿[65].24-表油菜素内酯(24-epibrassinolide, EBR)是BR家族中的一种.有报道发现,EBR可增加富铁幼苗根系高铁还原酶FRO的活性,却降低了缺铁幼苗中FRO的活性;进一步研究发现,EBR抑制缺铁诱导的乙烯产生,但促进富铁幼苗中乙烯的产生,说明BR可能通过与乙烯交互负调控FRO活性,从而参与植物对缺铁的响应[66]. ...
Brassinosteroids are involved in response of cucumber (Cucumis sativus) to iron deficiency
1
2012
... 乙烯的信号转导及功能受到泛素-蛋白酶体系统的调节[62].ACS作为乙烯生物合成的关键限速酶,其蛋白丰度受到ETO1(ethylene overproducing 1)及其同源蛋白EOL1(ETO1-like)和EOL2的调控[63].ETO1/EOLs是CULLIN3 E3泛素连接酶的组分,可以特异性识别2型ACS并通过26S蛋白酶体进行快速降解[63].14-3-3蛋白是真核生物中高度保守的二聚体蛋白,通过磷酸化介导与靶蛋白相互作用参与多种生物过程[64].在碳饥饿条件下,拟南芥出现叶片黄化衰老的表型,同时,碳饥饿会激活油菜素甾醇(brassinosteroid, BR)信号通路,由BR诱导的磷酸化的SINAT2(seven-in-absentia of A. thaliana 2)和ETO1/EOL2将14-3-3蛋白招募到复合物中,激活SINAT2与EOL2的降解,增加ACS5蛋白的丰度和自噬活性,最终促进乙烯生物合成以响应碳饥饿[65].24-表油菜素内酯(24-epibrassinolide, EBR)是BR家族中的一种.有报道发现,EBR可增加富铁幼苗根系高铁还原酶FRO的活性,却降低了缺铁幼苗中FRO的活性;进一步研究发现,EBR抑制缺铁诱导的乙烯产生,但促进富铁幼苗中乙烯的产生,说明BR可能通过与乙烯交互负调控FRO活性,从而参与植物对缺铁的响应[66]. ...
A new model involving ethylene, nitric oxide and Fe to explain the regulation of Fe-acquisition genes in strategy I plants
1
2011
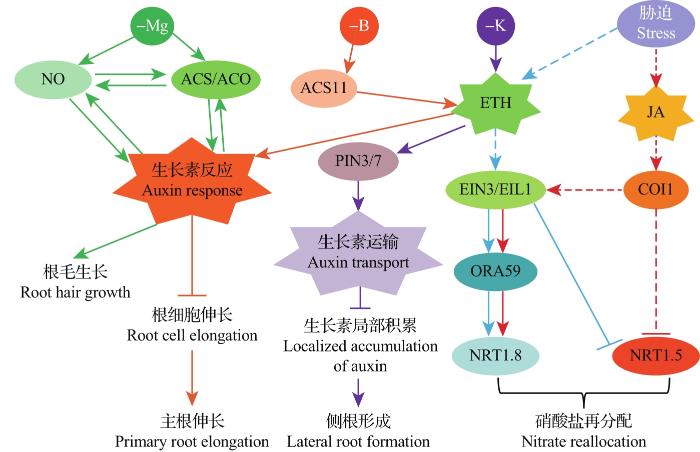
... 乙烯和一氧化氮(nitric oxide, NO)共同作用调节拟南芥缺铁和缺镁(Mg)响应.研究发现,在缺铁条件下,NO可以通过诱导乙烯合成基因表达来提高根系中乙烯的产量,而乙烯反过来可以提高根系中NO的产量,导致缺铁胁迫信号被放大,并最终产生一个激活信号来上调铁获取基因(FIT、bHLH38等)的表达[67].在缺镁条件下,乙烯和NO均通过诱导AUX1(AUXIN-RESISTANT 1)、PIN1和PIN2基因的表达,促进根系生长素水平的升高.反过来,生长素通过激活 ACO、ACS、硝酸还原酶和类NO合酶(NO synthase-like, NOS-L)的活性来刺激乙烯和NO的产生,从而形成ETH-NO-IAA的正反馈环路.此外,生长素作用于乙烯和NO信号的下游,调节缺镁诱导的根毛形态发生[68]. ...
Auxin acts downstream of ethylene and nitric oxide to regulate magnesium deficiency-induced root hair development in Arabidopsis thaliana
1
2018
... 乙烯和一氧化氮(nitric oxide, NO)共同作用调节拟南芥缺铁和缺镁(Mg)响应.研究发现,在缺铁条件下,NO可以通过诱导乙烯合成基因表达来提高根系中乙烯的产量,而乙烯反过来可以提高根系中NO的产量,导致缺铁胁迫信号被放大,并最终产生一个激活信号来上调铁获取基因(FIT、bHLH38等)的表达[67].在缺镁条件下,乙烯和NO均通过诱导AUX1(AUXIN-RESISTANT 1)、PIN1和PIN2基因的表达,促进根系生长素水平的升高.反过来,生长素通过激活 ACO、ACS、硝酸还原酶和类NO合酶(NO synthase-like, NOS-L)的活性来刺激乙烯和NO的产生,从而形成ETH-NO-IAA的正反馈环路.此外,生长素作用于乙烯和NO信号的下游,调节缺镁诱导的根毛形态发生[68]. ...
The TOR-EIN2 axis mediates nuclear signalling to modulate plant growth
1
2021
... 在信号转导层面,乙烯通过核心转录因子EIN3/EIL1参与调控缺铁、缺磷、缺硫等多种营养胁迫反应,例如:乙烯通过EIN3/EIL1与FIT互作,调控FRO2和IRT1表达;通过EIN3/EIL1与WRKY75互作,调控PHT1;5表达等.在其他营养缺乏条件下,乙烯是否也通过EIN3/EIL1这个关键节点来调控相关基因的表达以响应营养胁迫是一个值得探讨的问题.并且,EIN3/EIL1与特定营养信号通路中关键调控因子的互作可能是保证该信号通路特异性的重要方式.除了EIN3/EIL1,EIN2是乙烯信号通路的另一个关键蛋白.最新的研究表明,EIN2是雷帕霉素靶蛋白(target of rapamycin, TOR)激酶的一个新底物,TOR的功能被抑制可以促进EIN2全长蛋白进入细胞核,这种调控模式有别于经典的乙烯信号转导[69].其他营养胁迫信号是否也会以同样或类似的方式影响EIN2蛋白功能是另一个值得探讨的问题.此外,乙烯与多种激素信号存在交互作用并共同调控营养缺乏胁迫响应,但它们交叉互作的很多关键节点还不清楚.总之,乙烯与营养信号之间的相互作用有助于植物应对不断变化的营养环境,全面了解其作用及分子机制,将为作物养分高效利用的分子育种提供新策略. ...
Q & A: How do plants respond to ethylene and what is its importance?
1
2016
... 乙烯是迄今发现的唯一一种植物气体激素,在植物生长发育的多个方面发挥着重要功能,包括促进种子萌发、抑制植物根的伸长、促进植物根毛发育、调节花和叶片的衰老脱落等[1].乙烯是由甲硫氨酸(methionine, Met)的活化形式S-腺苷甲硫氨酸(S-adenosylmethionine, SAM)经ACC合酶(ACC synthase, ACS)转化为1-氨基环丙烷-1-羧酸(1-aminocyclopropane-1-carboxylic acid, ACC),再经ACC氧化酶(ACC oxidase, ACO)氧化所生成的[2].在信号转导过程中,首先是乙烯受体ETR1(ethylene response 1)、ERS1(ethylene response sensor 1)、ETR2、ERS2和EIN4(ethylene insensitive 4)以二聚体的形式通过亚铜离子结合乙烯而失去活性,从而使丝氨酸/苏氨酸蛋白激酶CTR1(constitutive triple response 1)失活,进而使未被CTR1磷酸化的膜蛋白EIN2的C末端(EIN2-C)得以被未知蛋白切割下来[3].其次,EIN2-C一方面通过核定位信号进入细胞核,并通过ENAP1(EIN2 nuclear associated protein 1)调控组蛋白H3K14和H3K23的乙酰化,进而使EIN3/EIL1(ethylene-insensitive 3-like 1)转录激活ERF1等下游靶基因表达,从而产生乙烯反应[4];另一方面则进入细胞质与EBF1/2(EIN3 binding F-box 1/2)mRNA的3非翻译区结合,并通过与多个P-body(processing body)元件相互作用,将其靶向至P-body,从而抑制EBF1和EBF2的翻译,最终促进EIN3/EIL1蛋白积累,触发乙烯引起的一系列生长发育响应[5]. ...
Accu-mulation and transport of 1-aminocyclopropane-1-carboxylic acid (ACC) in plants: current status, considerations for future research and agronomic applications
1
2017
... 乙烯是迄今发现的唯一一种植物气体激素,在植物生长发育的多个方面发挥着重要功能,包括促进种子萌发、抑制植物根的伸长、促进植物根毛发育、调节花和叶片的衰老脱落等[1].乙烯是由甲硫氨酸(methionine, Met)的活化形式S-腺苷甲硫氨酸(S-adenosylmethionine, SAM)经ACC合酶(ACC synthase, ACS)转化为1-氨基环丙烷-1-羧酸(1-aminocyclopropane-1-carboxylic acid, ACC),再经ACC氧化酶(ACC oxidase, ACO)氧化所生成的[2].在信号转导过程中,首先是乙烯受体ETR1(ethylene response 1)、ERS1(ethylene response sensor 1)、ETR2、ERS2和EIN4(ethylene insensitive 4)以二聚体的形式通过亚铜离子结合乙烯而失去活性,从而使丝氨酸/苏氨酸蛋白激酶CTR1(constitutive triple response 1)失活,进而使未被CTR1磷酸化的膜蛋白EIN2的C末端(EIN2-C)得以被未知蛋白切割下来[3].其次,EIN2-C一方面通过核定位信号进入细胞核,并通过ENAP1(EIN2 nuclear associated protein 1)调控组蛋白H3K14和H3K23的乙酰化,进而使EIN3/EIL1(ethylene-insensitive 3-like 1)转录激活ERF1等下游靶基因表达,从而产生乙烯反应[4];另一方面则进入细胞质与EBF1/2(EIN3 binding F-box 1/2)mRNA的3非翻译区结合,并通过与多个P-body(processing body)元件相互作用,将其靶向至P-body,从而抑制EBF1和EBF2的翻译,最终促进EIN3/EIL1蛋白积累,触发乙烯引起的一系列生长发育响应[5]. ...
Activation of ethylene signaling is mediated by nuclear translocation of the cleaved EIN2 carboxyl terminus
1
2012
... 乙烯是迄今发现的唯一一种植物气体激素,在植物生长发育的多个方面发挥着重要功能,包括促进种子萌发、抑制植物根的伸长、促进植物根毛发育、调节花和叶片的衰老脱落等[1].乙烯是由甲硫氨酸(methionine, Met)的活化形式S-腺苷甲硫氨酸(S-adenosylmethionine, SAM)经ACC合酶(ACC synthase, ACS)转化为1-氨基环丙烷-1-羧酸(1-aminocyclopropane-1-carboxylic acid, ACC),再经ACC氧化酶(ACC oxidase, ACO)氧化所生成的[2].在信号转导过程中,首先是乙烯受体ETR1(ethylene response 1)、ERS1(ethylene response sensor 1)、ETR2、ERS2和EIN4(ethylene insensitive 4)以二聚体的形式通过亚铜离子结合乙烯而失去活性,从而使丝氨酸/苏氨酸蛋白激酶CTR1(constitutive triple response 1)失活,进而使未被CTR1磷酸化的膜蛋白EIN2的C末端(EIN2-C)得以被未知蛋白切割下来[3].其次,EIN2-C一方面通过核定位信号进入细胞核,并通过ENAP1(EIN2 nuclear associated protein 1)调控组蛋白H3K14和H3K23的乙酰化,进而使EIN3/EIL1(ethylene-insensitive 3-like 1)转录激活ERF1等下游靶基因表达,从而产生乙烯反应[4];另一方面则进入细胞质与EBF1/2(EIN3 binding F-box 1/2)mRNA的3非翻译区结合,并通过与多个P-body(processing body)元件相互作用,将其靶向至P-body,从而抑制EBF1和EBF2的翻译,最终促进EIN3/EIL1蛋白积累,触发乙烯引起的一系列生长发育响应[5]. ...
EIN2 mediates direct regulation of histone acetylation in the ethylene response
1
2017
... 乙烯是迄今发现的唯一一种植物气体激素,在植物生长发育的多个方面发挥着重要功能,包括促进种子萌发、抑制植物根的伸长、促进植物根毛发育、调节花和叶片的衰老脱落等[1].乙烯是由甲硫氨酸(methionine, Met)的活化形式S-腺苷甲硫氨酸(S-adenosylmethionine, SAM)经ACC合酶(ACC synthase, ACS)转化为1-氨基环丙烷-1-羧酸(1-aminocyclopropane-1-carboxylic acid, ACC),再经ACC氧化酶(ACC oxidase, ACO)氧化所生成的[2].在信号转导过程中,首先是乙烯受体ETR1(ethylene response 1)、ERS1(ethylene response sensor 1)、ETR2、ERS2和EIN4(ethylene insensitive 4)以二聚体的形式通过亚铜离子结合乙烯而失去活性,从而使丝氨酸/苏氨酸蛋白激酶CTR1(constitutive triple response 1)失活,进而使未被CTR1磷酸化的膜蛋白EIN2的C末端(EIN2-C)得以被未知蛋白切割下来[3].其次,EIN2-C一方面通过核定位信号进入细胞核,并通过ENAP1(EIN2 nuclear associated protein 1)调控组蛋白H3K14和H3K23的乙酰化,进而使EIN3/EIL1(ethylene-insensitive 3-like 1)转录激活ERF1等下游靶基因表达,从而产生乙烯反应[4];另一方面则进入细胞质与EBF1/2(EIN3 binding F-box 1/2)mRNA的3非翻译区结合,并通过与多个P-body(processing body)元件相互作用,将其靶向至P-body,从而抑制EBF1和EBF2的翻译,最终促进EIN3/EIL1蛋白积累,触发乙烯引起的一系列生长发育响应[5]. ...
EIN2-directed translational regulation of ethylene signaling in Arabidopsis
1
2015
... 乙烯是迄今发现的唯一一种植物气体激素,在植物生长发育的多个方面发挥着重要功能,包括促进种子萌发、抑制植物根的伸长、促进植物根毛发育、调节花和叶片的衰老脱落等[1].乙烯是由甲硫氨酸(methionine, Met)的活化形式S-腺苷甲硫氨酸(S-adenosylmethionine, SAM)经ACC合酶(ACC synthase, ACS)转化为1-氨基环丙烷-1-羧酸(1-aminocyclopropane-1-carboxylic acid, ACC),再经ACC氧化酶(ACC oxidase, ACO)氧化所生成的[2].在信号转导过程中,首先是乙烯受体ETR1(ethylene response 1)、ERS1(ethylene response sensor 1)、ETR2、ERS2和EIN4(ethylene insensitive 4)以二聚体的形式通过亚铜离子结合乙烯而失去活性,从而使丝氨酸/苏氨酸蛋白激酶CTR1(constitutive triple response 1)失活,进而使未被CTR1磷酸化的膜蛋白EIN2的C末端(EIN2-C)得以被未知蛋白切割下来[3].其次,EIN2-C一方面通过核定位信号进入细胞核,并通过ENAP1(EIN2 nuclear associated protein 1)调控组蛋白H3K14和H3K23的乙酰化,进而使EIN3/EIL1(ethylene-insensitive 3-like 1)转录激活ERF1等下游靶基因表达,从而产生乙烯反应[4];另一方面则进入细胞质与EBF1/2(EIN3 binding F-box 1/2)mRNA的3非翻译区结合,并通过与多个P-body(processing body)元件相互作用,将其靶向至P-body,从而抑制EBF1和EBF2的翻译,最终促进EIN3/EIL1蛋白积累,触发乙烯引起的一系列生长发育响应[5]. ...
Iron: nutritious, noxious, and not readily available
1
1994
... 铁是植物必需的微量元素,尽管在地壳中含量丰富,但在碱性或石灰性土壤中,铁的有效性很低,不能满足植物正常生长发育所需,导致植物缺铁,出现叶片失绿症状,抑制生长发育,降低植物的产量和品质[6].植物缺铁时,会诱导根的生理和形态发生变化,以提高铁的活化、吸收和转运效率,称为“缺铁响应”[7].缺铁响应调控是乙烯参与养分吸收利用过程最早得到报道且研究最为深入的.缺铁可以促进乙烯的生物合成和信号转导,例如,缺铁诱导拟南芥(Arabidopsis thaliana)根中乙烯合成基因(如AtSAM1、AtSAM2、AtACS4、AtACS6、AtACS9、AtACO1、AtACO2)和信号转导基因(如AtETR1、AtCTR1、AtEIN2、AtEIN3、AtEIL1、AtEIL3)的表达[8],而乙烯又可以调控根系的发生、发育及其功能. ...
Ethylene and nitric oxide involvement in the regulation of Fe and P deficiency responses in dicotyledonous plants
1
2021
... 铁是植物必需的微量元素,尽管在地壳中含量丰富,但在碱性或石灰性土壤中,铁的有效性很低,不能满足植物正常生长发育所需,导致植物缺铁,出现叶片失绿症状,抑制生长发育,降低植物的产量和品质[6].植物缺铁时,会诱导根的生理和形态发生变化,以提高铁的活化、吸收和转运效率,称为“缺铁响应”[7].缺铁响应调控是乙烯参与养分吸收利用过程最早得到报道且研究最为深入的.缺铁可以促进乙烯的生物合成和信号转导,例如,缺铁诱导拟南芥(Arabidopsis thaliana)根中乙烯合成基因(如AtSAM1、AtSAM2、AtACS4、AtACS6、AtACS9、AtACO1、AtACO2)和信号转导基因(如AtETR1、AtCTR1、AtEIN2、AtEIN3、AtEIL1、AtEIL3)的表达[8],而乙烯又可以调控根系的发生、发育及其功能. ...
Ethylene and nitric oxide involvement in the up-regulation of key genes related to iron acquisition and homeostasis in Arabidopsis
2
2010
... 铁是植物必需的微量元素,尽管在地壳中含量丰富,但在碱性或石灰性土壤中,铁的有效性很低,不能满足植物正常生长发育所需,导致植物缺铁,出现叶片失绿症状,抑制生长发育,降低植物的产量和品质[6].植物缺铁时,会诱导根的生理和形态发生变化,以提高铁的活化、吸收和转运效率,称为“缺铁响应”[7].缺铁响应调控是乙烯参与养分吸收利用过程最早得到报道且研究最为深入的.缺铁可以促进乙烯的生物合成和信号转导,例如,缺铁诱导拟南芥(Arabidopsis thaliana)根中乙烯合成基因(如AtSAM1、AtSAM2、AtACS4、AtACS6、AtACS9、AtACO1、AtACO2)和信号转导基因(如AtETR1、AtCTR1、AtEIN2、AtEIN3、AtEIL1、AtEIL3)的表达[8],而乙烯又可以调控根系的发生、发育及其功能. ...
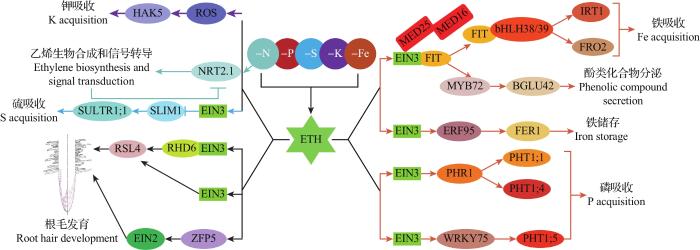
... 在分子生理水平上,乙烯可以通过调节铁吸收、活化和运输相关的调控元件来影响植物的缺铁响应.FIT(FER-like iron deficiency-induced trans-cription factor)/AtbHLH29属于碱性螺旋-环-螺旋(basic helix-loop-helix, bHLH)蛋白家族转录因子,其通过与AtbHLH38和AtbHLH39相互作用形成异源二聚体,调控高铁还原酶基因FRO2(FERRIC REDUCTION OXIDASE 2)和铁转运基因IRT1(IRON-REGULATED TRANSPORTER 1)的表达,在缺铁响应中发挥着核心调控作用[9-11].乙烯可上调AtFIT(或其番茄同源物SlFER)、AtbHLH38和AtbHLH39的表达[12],其机制是乙烯通过信号转导通路中的核心转录因子AtEIN3/AtEIL1一方面与AtFIT相互作用,阻止它被蛋白酶体降解,从而影响其转录后的稳定性[13],另一方面通过与AtMED16(mediator subunit 16 in A. thaliana)和AtMED25相互作用形成一个复合体来参与AtFIT的转录,调控下游基因AtIRT1和AtFRO2的表达[14].可见,乙烯是在转录和转录后水平上控制AtFIT水平的重要信号.除了提高铁吸收能力,乙烯在缺铁条件下还可以通过调节酚类化合物的分泌促进铁的活化.研究发现,AtEIN3/AtEIL1/AtFIT复合物通过调控AtMYB72的转录间接调控β-葡萄糖苷酶AtBGLU42的表达[15],而AtBGLU42对酚类化合物的分泌非常重要[16].此外,在缺铁条件下,乙烯还通过参与激活AtFRD3和AtNASs等基因的表达[8],促进植物体内铁的运输和分配[17]. ...
Molecular mechanisms governing Arabidopsis iron uptake
1
2015
... 在分子生理水平上,乙烯可以通过调节铁吸收、活化和运输相关的调控元件来影响植物的缺铁响应.FIT(FER-like iron deficiency-induced trans-cription factor)/AtbHLH29属于碱性螺旋-环-螺旋(basic helix-loop-helix, bHLH)蛋白家族转录因子,其通过与AtbHLH38和AtbHLH39相互作用形成异源二聚体,调控高铁还原酶基因FRO2(FERRIC REDUCTION OXIDASE 2)和铁转运基因IRT1(IRON-REGULATED TRANSPORTER 1)的表达,在缺铁响应中发挥着核心调控作用[9-11].乙烯可上调AtFIT(或其番茄同源物SlFER)、AtbHLH38和AtbHLH39的表达[12],其机制是乙烯通过信号转导通路中的核心转录因子AtEIN3/AtEIL1一方面与AtFIT相互作用,阻止它被蛋白酶体降解,从而影响其转录后的稳定性[13],另一方面通过与AtMED16(mediator subunit 16 in A. thaliana)和AtMED25相互作用形成一个复合体来参与AtFIT的转录,调控下游基因AtIRT1和AtFRO2的表达[14].可见,乙烯是在转录和转录后水平上控制AtFIT水平的重要信号.除了提高铁吸收能力,乙烯在缺铁条件下还可以通过调节酚类化合物的分泌促进铁的活化.研究发现,AtEIN3/AtEIL1/AtFIT复合物通过调控AtMYB72的转录间接调控β-葡萄糖苷酶AtBGLU42的表达[15],而AtBGLU42对酚类化合物的分泌非常重要[16].此外,在缺铁条件下,乙烯还通过参与激活AtFRD3和AtNASs等基因的表达[8],促进植物体内铁的运输和分配[17]. ...
The transcriptional control of iron homeostasis in plants: a tale of bHLH transcription factors?
0
2019
FIT-dependent and -independent gene signatures
1
2020
... 在分子生理水平上,乙烯可以通过调节铁吸收、活化和运输相关的调控元件来影响植物的缺铁响应.FIT(FER-like iron deficiency-induced trans-cription factor)/AtbHLH29属于碱性螺旋-环-螺旋(basic helix-loop-helix, bHLH)蛋白家族转录因子,其通过与AtbHLH38和AtbHLH39相互作用形成异源二聚体,调控高铁还原酶基因FRO2(FERRIC REDUCTION OXIDASE 2)和铁转运基因IRT1(IRON-REGULATED TRANSPORTER 1)的表达,在缺铁响应中发挥着核心调控作用[9-11].乙烯可上调AtFIT(或其番茄同源物SlFER)、AtbHLH38和AtbHLH39的表达[12],其机制是乙烯通过信号转导通路中的核心转录因子AtEIN3/AtEIL1一方面与AtFIT相互作用,阻止它被蛋白酶体降解,从而影响其转录后的稳定性[13],另一方面通过与AtMED16(mediator subunit 16 in A. thaliana)和AtMED25相互作用形成一个复合体来参与AtFIT的转录,调控下游基因AtIRT1和AtFRO2的表达[14].可见,乙烯是在转录和转录后水平上控制AtFIT水平的重要信号.除了提高铁吸收能力,乙烯在缺铁条件下还可以通过调节酚类化合物的分泌促进铁的活化.研究发现,AtEIN3/AtEIL1/AtFIT复合物通过调控AtMYB72的转录间接调控β-葡萄糖苷酶AtBGLU42的表达[15],而AtBGLU42对酚类化合物的分泌非常重要[16].此外,在缺铁条件下,乙烯还通过参与激活AtFRD3和AtNASs等基因的表达[8],促进植物体内铁的运输和分配[17]. ...
Hypoxia and bicarbonate could limit the expression of iron acquisition genes in strategy I plants by affecting ethylene synthesis and signaling in different ways
1
2014
... 在分子生理水平上,乙烯可以通过调节铁吸收、活化和运输相关的调控元件来影响植物的缺铁响应.FIT(FER-like iron deficiency-induced trans-cription factor)/AtbHLH29属于碱性螺旋-环-螺旋(basic helix-loop-helix, bHLH)蛋白家族转录因子,其通过与AtbHLH38和AtbHLH39相互作用形成异源二聚体,调控高铁还原酶基因FRO2(FERRIC REDUCTION OXIDASE 2)和铁转运基因IRT1(IRON-REGULATED TRANSPORTER 1)的表达,在缺铁响应中发挥着核心调控作用[9-11].乙烯可上调AtFIT(或其番茄同源物SlFER)、AtbHLH38和AtbHLH39的表达[12],其机制是乙烯通过信号转导通路中的核心转录因子AtEIN3/AtEIL1一方面与AtFIT相互作用,阻止它被蛋白酶体降解,从而影响其转录后的稳定性[13],另一方面通过与AtMED16(mediator subunit 16 in A. thaliana)和AtMED25相互作用形成一个复合体来参与AtFIT的转录,调控下游基因AtIRT1和AtFRO2的表达[14].可见,乙烯是在转录和转录后水平上控制AtFIT水平的重要信号.除了提高铁吸收能力,乙烯在缺铁条件下还可以通过调节酚类化合物的分泌促进铁的活化.研究发现,AtEIN3/AtEIL1/AtFIT复合物通过调控AtMYB72的转录间接调控β-葡萄糖苷酶AtBGLU42的表达[15],而AtBGLU42对酚类化合物的分泌非常重要[16].此外,在缺铁条件下,乙烯还通过参与激活AtFRD3和AtNASs等基因的表达[8],促进植物体内铁的运输和分配[17]. ...
Interaction between the bHLH transcription factor FIT and ETHYLENE INSENSITIVE 3/ETHYLENE INSENSITIVE 3-LIKE 1 reveals molecular linkage between the regulation of iron acquisition and ethylene signaling in Arabidopsis
1
2011
... 在分子生理水平上,乙烯可以通过调节铁吸收、活化和运输相关的调控元件来影响植物的缺铁响应.FIT(FER-like iron deficiency-induced trans-cription factor)/AtbHLH29属于碱性螺旋-环-螺旋(basic helix-loop-helix, bHLH)蛋白家族转录因子,其通过与AtbHLH38和AtbHLH39相互作用形成异源二聚体,调控高铁还原酶基因FRO2(FERRIC REDUCTION OXIDASE 2)和铁转运基因IRT1(IRON-REGULATED TRANSPORTER 1)的表达,在缺铁响应中发挥着核心调控作用[9-11].乙烯可上调AtFIT(或其番茄同源物SlFER)、AtbHLH38和AtbHLH39的表达[12],其机制是乙烯通过信号转导通路中的核心转录因子AtEIN3/AtEIL1一方面与AtFIT相互作用,阻止它被蛋白酶体降解,从而影响其转录后的稳定性[13],另一方面通过与AtMED16(mediator subunit 16 in A. thaliana)和AtMED25相互作用形成一个复合体来参与AtFIT的转录,调控下游基因AtIRT1和AtFRO2的表达[14].可见,乙烯是在转录和转录后水平上控制AtFIT水平的重要信号.除了提高铁吸收能力,乙烯在缺铁条件下还可以通过调节酚类化合物的分泌促进铁的活化.研究发现,AtEIN3/AtEIL1/AtFIT复合物通过调控AtMYB72的转录间接调控β-葡萄糖苷酶AtBGLU42的表达[15],而AtBGLU42对酚类化合物的分泌非常重要[16].此外,在缺铁条件下,乙烯还通过参与激活AtFRD3和AtNASs等基因的表达[8],促进植物体内铁的运输和分配[17]. ...
The Arabidopsis mediator subunit MED16 regulates iron homeostasis by associating with EIN3/EIL1 through subunit MED25
1
2014
... 在分子生理水平上,乙烯可以通过调节铁吸收、活化和运输相关的调控元件来影响植物的缺铁响应.FIT(FER-like iron deficiency-induced trans-cription factor)/AtbHLH29属于碱性螺旋-环-螺旋(basic helix-loop-helix, bHLH)蛋白家族转录因子,其通过与AtbHLH38和AtbHLH39相互作用形成异源二聚体,调控高铁还原酶基因FRO2(FERRIC REDUCTION OXIDASE 2)和铁转运基因IRT1(IRON-REGULATED TRANSPORTER 1)的表达,在缺铁响应中发挥着核心调控作用[9-11].乙烯可上调AtFIT(或其番茄同源物SlFER)、AtbHLH38和AtbHLH39的表达[12],其机制是乙烯通过信号转导通路中的核心转录因子AtEIN3/AtEIL1一方面与AtFIT相互作用,阻止它被蛋白酶体降解,从而影响其转录后的稳定性[13],另一方面通过与AtMED16(mediator subunit 16 in A. thaliana)和AtMED25相互作用形成一个复合体来参与AtFIT的转录,调控下游基因AtIRT1和AtFRO2的表达[14].可见,乙烯是在转录和转录后水平上控制AtFIT水平的重要信号.除了提高铁吸收能力,乙烯在缺铁条件下还可以通过调节酚类化合物的分泌促进铁的活化.研究发现,AtEIN3/AtEIL1/AtFIT复合物通过调控AtMYB72的转录间接调控β-葡萄糖苷酶AtBGLU42的表达[15],而AtBGLU42对酚类化合物的分泌非常重要[16].此外,在缺铁条件下,乙烯还通过参与激活AtFRD3和AtNASs等基因的表达[8],促进植物体内铁的运输和分配[17]. ...
New routes for plant iron mining
1
2017
... 在分子生理水平上,乙烯可以通过调节铁吸收、活化和运输相关的调控元件来影响植物的缺铁响应.FIT(FER-like iron deficiency-induced trans-cription factor)/AtbHLH29属于碱性螺旋-环-螺旋(basic helix-loop-helix, bHLH)蛋白家族转录因子,其通过与AtbHLH38和AtbHLH39相互作用形成异源二聚体,调控高铁还原酶基因FRO2(FERRIC REDUCTION OXIDASE 2)和铁转运基因IRT1(IRON-REGULATED TRANSPORTER 1)的表达,在缺铁响应中发挥着核心调控作用[9-11].乙烯可上调AtFIT(或其番茄同源物SlFER)、AtbHLH38和AtbHLH39的表达[12],其机制是乙烯通过信号转导通路中的核心转录因子AtEIN3/AtEIL1一方面与AtFIT相互作用,阻止它被蛋白酶体降解,从而影响其转录后的稳定性[13],另一方面通过与AtMED16(mediator subunit 16 in A. thaliana)和AtMED25相互作用形成一个复合体来参与AtFIT的转录,调控下游基因AtIRT1和AtFRO2的表达[14].可见,乙烯是在转录和转录后水平上控制AtFIT水平的重要信号.除了提高铁吸收能力,乙烯在缺铁条件下还可以通过调节酚类化合物的分泌促进铁的活化.研究发现,AtEIN3/AtEIL1/AtFIT复合物通过调控AtMYB72的转录间接调控β-葡萄糖苷酶AtBGLU42的表达[15],而AtBGLU42对酚类化合物的分泌非常重要[16].此外,在缺铁条件下,乙烯还通过参与激活AtFRD3和AtNASs等基因的表达[8],促进植物体内铁的运输和分配[17]. ...
Beta-glucosidase BGLU42 is a MYB72-dependent key regulator of rhizobacteria-induced systemic resistance and modulates iron deficiency responses in Arabidopsis roots
1
2014
... 在分子生理水平上,乙烯可以通过调节铁吸收、活化和运输相关的调控元件来影响植物的缺铁响应.FIT(FER-like iron deficiency-induced trans-cription factor)/AtbHLH29属于碱性螺旋-环-螺旋(basic helix-loop-helix, bHLH)蛋白家族转录因子,其通过与AtbHLH38和AtbHLH39相互作用形成异源二聚体,调控高铁还原酶基因FRO2(FERRIC REDUCTION OXIDASE 2)和铁转运基因IRT1(IRON-REGULATED TRANSPORTER 1)的表达,在缺铁响应中发挥着核心调控作用[9-11].乙烯可上调AtFIT(或其番茄同源物SlFER)、AtbHLH38和AtbHLH39的表达[12],其机制是乙烯通过信号转导通路中的核心转录因子AtEIN3/AtEIL1一方面与AtFIT相互作用,阻止它被蛋白酶体降解,从而影响其转录后的稳定性[13],另一方面通过与AtMED16(mediator subunit 16 in A. thaliana)和AtMED25相互作用形成一个复合体来参与AtFIT的转录,调控下游基因AtIRT1和AtFRO2的表达[14].可见,乙烯是在转录和转录后水平上控制AtFIT水平的重要信号.除了提高铁吸收能力,乙烯在缺铁条件下还可以通过调节酚类化合物的分泌促进铁的活化.研究发现,AtEIN3/AtEIL1/AtFIT复合物通过调控AtMYB72的转录间接调控β-葡萄糖苷酶AtBGLU42的表达[15],而AtBGLU42对酚类化合物的分泌非常重要[16].此外,在缺铁条件下,乙烯还通过参与激活AtFRD3和AtNASs等基因的表达[8],促进植物体内铁的运输和分配[17]. ...
Ethylene participates in the regulation of Fe deficiency responses in strategy I plants and in rice
2
2015
... 在分子生理水平上,乙烯可以通过调节铁吸收、活化和运输相关的调控元件来影响植物的缺铁响应.FIT(FER-like iron deficiency-induced trans-cription factor)/AtbHLH29属于碱性螺旋-环-螺旋(basic helix-loop-helix, bHLH)蛋白家族转录因子,其通过与AtbHLH38和AtbHLH39相互作用形成异源二聚体,调控高铁还原酶基因FRO2(FERRIC REDUCTION OXIDASE 2)和铁转运基因IRT1(IRON-REGULATED TRANSPORTER 1)的表达,在缺铁响应中发挥着核心调控作用[9-11].乙烯可上调AtFIT(或其番茄同源物SlFER)、AtbHLH38和AtbHLH39的表达[12],其机制是乙烯通过信号转导通路中的核心转录因子AtEIN3/AtEIL1一方面与AtFIT相互作用,阻止它被蛋白酶体降解,从而影响其转录后的稳定性[13],另一方面通过与AtMED16(mediator subunit 16 in A. thaliana)和AtMED25相互作用形成一个复合体来参与AtFIT的转录,调控下游基因AtIRT1和AtFRO2的表达[14].可见,乙烯是在转录和转录后水平上控制AtFIT水平的重要信号.除了提高铁吸收能力,乙烯在缺铁条件下还可以通过调节酚类化合物的分泌促进铁的活化.研究发现,AtEIN3/AtEIL1/AtFIT复合物通过调控AtMYB72的转录间接调控β-葡萄糖苷酶AtBGLU42的表达[15],而AtBGLU42对酚类化合物的分泌非常重要[16].此外,在缺铁条件下,乙烯还通过参与激活AtFRD3和AtNASs等基因的表达[8],促进植物体内铁的运输和分配[17]. ...
... 在形态发育水平上,乙烯直接参与缺铁条件下植物根毛、根表皮转移细胞和排根的发育以及主根变短和侧根数量增加等表型的形成[17-18].研究发现,EIN3/EIL1转录因子通过与RHD6(root hair defective 6)相互作用激活下游RSL4基因的表达,调控根毛发育[19].此外,乙烯通过诱导额外的根毛和转移细胞的形成,参与调控缺铁条件下根表皮细胞命运的适应性变化[20]. ...
Ethylene and the regulation of physiological and morphological responses to nutrient deficiencies
1
2015
... 在形态发育水平上,乙烯直接参与缺铁条件下植物根毛、根表皮转移细胞和排根的发育以及主根变短和侧根数量增加等表型的形成[17-18].研究发现,EIN3/EIL1转录因子通过与RHD6(root hair defective 6)相互作用激活下游RSL4基因的表达,调控根毛发育[19].此外,乙烯通过诱导额外的根毛和转移细胞的形成,参与调控缺铁条件下根表皮细胞命运的适应性变化[20]. ...
Ethylene promotes root hair growth through coordinated EIN3/EIL1 and RHD6/RSL1 activity in Arabidopsis
2
2017
... 在形态发育水平上,乙烯直接参与缺铁条件下植物根毛、根表皮转移细胞和排根的发育以及主根变短和侧根数量增加等表型的形成[17-18].研究发现,EIN3/EIL1转录因子通过与RHD6(root hair defective 6)相互作用激活下游RSL4基因的表达,调控根毛发育[19].此外,乙烯通过诱导额外的根毛和转移细胞的形成,参与调控缺铁条件下根表皮细胞命运的适应性变化[20]. ...
... 在形态发育水平上,植物可以通过重塑根系构型应对缺磷胁迫,包括抑制主根生长、刺激侧根发育与伸长、增加根毛数量、促进排根形成等[37].施用Ag+或AVG可以抑制低磷诱导的主根分生组织衰竭,表明乙烯参与了缺磷导致的主根生长抑制过程[38].研究发现,GmETO1(ethylene-overproduction protein 1 in G. max)通过调节乙烯合成和下游相关基因的表达,影响大豆毛状根的增殖和伸长,提高磷的吸收利用率,进而增强大豆对低磷胁迫的耐受性[39].缺磷诱导EIN3蛋白积累,而EIN3一方面直接调控RSL4(ROOT HAIR DEFECTIVE 6-LIKE 4)的表达[19],另一方面通过结合到一组调节细胞壁修饰的基因,如RHS19、EXT14的启动子上,激活它们的转录,进而促进根毛的发育[40]. ...
From faith to fate: ethylene signaling in morphogenic responses to P and Fe deficiency
1
2001
... 在形态发育水平上,乙烯直接参与缺铁条件下植物根毛、根表皮转移细胞和排根的发育以及主根变短和侧根数量增加等表型的形成[17-18].研究发现,EIN3/EIL1转录因子通过与RHD6(root hair defective 6)相互作用激活下游RSL4基因的表达,调控根毛发育[19].此外,乙烯通过诱导额外的根毛和转移细胞的形成,参与调控缺铁条件下根表皮细胞命运的适应性变化[20]. ...
Ethylene promotes seed iron storage during Arabidopsis seed maturation via ERF95 transcription factor
1
2020
... 除了上述作用,最新的研究表明,乙烯还参与调控种子铁储存过程.乙烯在拟南芥种子发育中后期合成量增加,其通过EIN3调控乙烯响应因子ERF95的表达,从而促进铁蛋白基因AtFer1的转录,而铁蛋白的累积不仅增加种子中铁的积累,而且可以缓冲种子铁储存过程中因铁过量引起的毒害作用,从而帮助更多的铁进入胚细胞液泡存储,以确保细胞内的铁稳态和子代幼苗早期的铁营养[21]. ...
Decreased ethylene biosynthesis, and induction of aerenchyma, by nitrogen- or phosphate-starvation in adventitious roots of Zea mays L.
2
1989
... 磷是植物正常生长发育和产量形成所需的大量元素之一,主要以无机磷酸根(Pi,H2PO4-)的形式被植物吸收.缺磷会改变植物乙烯的生物合成和信号转导.但缺磷对乙烯合成的影响因作物种类不同而异,如,玉米(Zea mays)[22]和番茄[23]中的乙烯含量降低,而普通大豆[Glycine max (Linn.) Merr.]根以及白羽扇豆(Lupinus albus)、黄花苜蓿(Medicago falcata L.)中的乙烯含量则会增加[24].在拟南芥中,缺磷能增强AtACS2、AtACS4和AtACS6的表达[25],并使编码乙烯响应转录因子ERF1和ERF2等基因的表达上调[26].研究表明,乙烯响应基因受缺磷的诱导表达在很大程度上依赖于缺磷响应核心调控因子PHR1(phosphate starvation response 1)及其同源蛋白AtPHL1[27].在phr1 phl1双突变体中,至少有8个AP2/ERF基因的表达相比于野生型下调[28].此外,缺磷也会改变植物对乙烯的敏感性.缺磷可增强玉米幼苗不定根[29]、菜豆(Phaseolus vulgaris Linn.)基根[30]和白三叶(Trifolium repens L.)侧根[31]对乙烯的敏感性,推测可能是因为缺磷改变了乙烯信号转导相关基因的表达. ...
... 硝态氮(NO3--N)和铵态氮(NH4+-N)是植物吸收的2种主要的无机氮源.除了作为营养物质,NO3--N和NH4+-N还能作为信号物质影响植物的生长发育[51].缺氮导致植物老叶变黄和生长发育迟缓[52].此外,缺氮会减少玉米根系乙烯的产生[22],但会增加根对乙烯的敏感性并诱导通气组织的形成[29],说明乙烯与氮素营养密切相关. ...
Ethylene insensitivity impedes a subset of responses to phosphorus deficiency in tomato and petunia
1
2008
... 磷是植物正常生长发育和产量形成所需的大量元素之一,主要以无机磷酸根(Pi,H2PO4-)的形式被植物吸收.缺磷会改变植物乙烯的生物合成和信号转导.但缺磷对乙烯合成的影响因作物种类不同而异,如,玉米(Zea mays)[22]和番茄[23]中的乙烯含量降低,而普通大豆[Glycine max (Linn.) Merr.]根以及白羽扇豆(Lupinus albus)、黄花苜蓿(Medicago falcata L.)中的乙烯含量则会增加[24].在拟南芥中,缺磷能增强AtACS2、AtACS4和AtACS6的表达[25],并使编码乙烯响应转录因子ERF1和ERF2等基因的表达上调[26].研究表明,乙烯响应基因受缺磷的诱导表达在很大程度上依赖于缺磷响应核心调控因子PHR1(phosphate starvation response 1)及其同源蛋白AtPHL1[27].在phr1 phl1双突变体中,至少有8个AP2/ERF基因的表达相比于野生型下调[28].此外,缺磷也会改变植物对乙烯的敏感性.缺磷可增强玉米幼苗不定根[29]、菜豆(Phaseolus vulgaris Linn.)基根[30]和白三叶(Trifolium repens L.)侧根[31]对乙烯的敏感性,推测可能是因为缺磷改变了乙烯信号转导相关基因的表达. ...
Phosphorus deficiency-induced reduction in root hydraulic conductivity in Medicago falcata is associated with ethylene production
1
2009
... 磷是植物正常生长发育和产量形成所需的大量元素之一,主要以无机磷酸根(Pi,H2PO4-)的形式被植物吸收.缺磷会改变植物乙烯的生物合成和信号转导.但缺磷对乙烯合成的影响因作物种类不同而异,如,玉米(Zea mays)[22]和番茄[23]中的乙烯含量降低,而普通大豆[Glycine max (Linn.) Merr.]根以及白羽扇豆(Lupinus albus)、黄花苜蓿(Medicago falcata L.)中的乙烯含量则会增加[24].在拟南芥中,缺磷能增强AtACS2、AtACS4和AtACS6的表达[25],并使编码乙烯响应转录因子ERF1和ERF2等基因的表达上调[26].研究表明,乙烯响应基因受缺磷的诱导表达在很大程度上依赖于缺磷响应核心调控因子PHR1(phosphate starvation response 1)及其同源蛋白AtPHL1[27].在phr1 phl1双突变体中,至少有8个AP2/ERF基因的表达相比于野生型下调[28].此外,缺磷也会改变植物对乙烯的敏感性.缺磷可增强玉米幼苗不定根[29]、菜豆(Phaseolus vulgaris Linn.)基根[30]和白三叶(Trifolium repens L.)侧根[31]对乙烯的敏感性,推测可能是因为缺磷改变了乙烯信号转导相关基因的表达. ...
Ethylene signalling is involved in regulation of phosphate starvation-induced gene expression and production of acid phosphatases and anthocyanin in Arabidopsis
6
2011
... 磷是植物正常生长发育和产量形成所需的大量元素之一,主要以无机磷酸根(Pi,H2PO4-)的形式被植物吸收.缺磷会改变植物乙烯的生物合成和信号转导.但缺磷对乙烯合成的影响因作物种类不同而异,如,玉米(Zea mays)[22]和番茄[23]中的乙烯含量降低,而普通大豆[Glycine max (Linn.) Merr.]根以及白羽扇豆(Lupinus albus)、黄花苜蓿(Medicago falcata L.)中的乙烯含量则会增加[24].在拟南芥中,缺磷能增强AtACS2、AtACS4和AtACS6的表达[25],并使编码乙烯响应转录因子ERF1和ERF2等基因的表达上调[26].研究表明,乙烯响应基因受缺磷的诱导表达在很大程度上依赖于缺磷响应核心调控因子PHR1(phosphate starvation response 1)及其同源蛋白AtPHL1[27].在phr1 phl1双突变体中,至少有8个AP2/ERF基因的表达相比于野生型下调[28].此外,缺磷也会改变植物对乙烯的敏感性.缺磷可增强玉米幼苗不定根[29]、菜豆(Phaseolus vulgaris Linn.)基根[30]和白三叶(Trifolium repens L.)侧根[31]对乙烯的敏感性,推测可能是因为缺磷改变了乙烯信号转导相关基因的表达. ...
... 在分子生理水平上,乙烯通过调节磷吸收、活化和再利用相关基因来调控植物缺磷响应.磷转运蛋白基因PT1(PHT1;1)和PT2(PHT1;4)的表达水平在拟南芥乙烯不敏感突变体etr1和ein2-5中显著降低,而在拟南芥组成型乙烯反应突变体hps2(hypersensitive to Pi starvation 2,ctr1同源物)中显著增加[25].已有研究表明,ACC可以诱导缺磷响应核心调控因子PHR1的表达,后者调控一系列磷吸收、转运和饥饿应答相关基因的表达[32].EIN3/EIL1通过调控转录因子WRKY75激活PHT1;5的表达[33].在其他植物中也有类似的发现,例如在苜蓿中,ACC在磷充足的条件下诱导磷转运蛋白基因MfPT1和MfPT5的表达,而乙烯生物合成抑制剂氨基乙氧基乙烯基甘氨酸(aminoethoxyvinylglycine, AVG)和Co2+均阻止了这些基因的低磷诱导表达[34].在低磷胁迫下,植物通过分泌酸性磷酸酶(acid phosphatase, APase)来活化土壤中的磷,以提高对有机磷的利用率.研究发现,拟南芥hps2突变体根表面的APase活性明显增加,而其ein2突变体的APase活性则显著降低,表明乙烯是APase活性的正调控因子[25].此外,花青素累积是缺磷的标志性症状,在低磷条件下,拟南芥hps2、hps3和hps4突变体中花青素的积累量低于野生型[25,35],但在ein2突变体中花青素含量则明显增加,说明乙烯是磷饥饿诱导花青素积累的负调控因子[25].hps2、hps3等突变体中花青素的减少可能是由于激活的乙烯信号通路缓解了低磷胁迫[25].在体内磷再利用方面,乙烯可促进拟南芥细胞壁磷的再利用,NAC转录因子ANAC044作为转录抑制因子,通过下调ACS6和ACO1的表达来抑制缺磷诱导的乙烯生物合成,从而抑制细胞壁磷的再利用[36]. ...
... [25].此外,花青素累积是缺磷的标志性症状,在低磷条件下,拟南芥hps2、hps3和hps4突变体中花青素的积累量低于野生型[25,35],但在ein2突变体中花青素含量则明显增加,说明乙烯是磷饥饿诱导花青素积累的负调控因子[25].hps2、hps3等突变体中花青素的减少可能是由于激活的乙烯信号通路缓解了低磷胁迫[25].在体内磷再利用方面,乙烯可促进拟南芥细胞壁磷的再利用,NAC转录因子ANAC044作为转录抑制因子,通过下调ACS6和ACO1的表达来抑制缺磷诱导的乙烯生物合成,从而抑制细胞壁磷的再利用[36]. ...
... [25,35],但在ein2突变体中花青素含量则明显增加,说明乙烯是磷饥饿诱导花青素积累的负调控因子[25].hps2、hps3等突变体中花青素的减少可能是由于激活的乙烯信号通路缓解了低磷胁迫[25].在体内磷再利用方面,乙烯可促进拟南芥细胞壁磷的再利用,NAC转录因子ANAC044作为转录抑制因子,通过下调ACS6和ACO1的表达来抑制缺磷诱导的乙烯生物合成,从而抑制细胞壁磷的再利用[36]. ...
... [25].hps2、hps3等突变体中花青素的减少可能是由于激活的乙烯信号通路缓解了低磷胁迫[25].在体内磷再利用方面,乙烯可促进拟南芥细胞壁磷的再利用,NAC转录因子ANAC044作为转录抑制因子,通过下调ACS6和ACO1的表达来抑制缺磷诱导的乙烯生物合成,从而抑制细胞壁磷的再利用[36]. ...
... [25].在体内磷再利用方面,乙烯可促进拟南芥细胞壁磷的再利用,NAC转录因子ANAC044作为转录抑制因子,通过下调ACS6和ACO1的表达来抑制缺磷诱导的乙烯生物合成,从而抑制细胞壁磷的再利用[36]. ...
Dissection of local and systemic transcriptional responses to phosphate starvation in Arabidopsis
1
2010
... 磷是植物正常生长发育和产量形成所需的大量元素之一,主要以无机磷酸根(Pi,H2PO4-)的形式被植物吸收.缺磷会改变植物乙烯的生物合成和信号转导.但缺磷对乙烯合成的影响因作物种类不同而异,如,玉米(Zea mays)[22]和番茄[23]中的乙烯含量降低,而普通大豆[Glycine max (Linn.) Merr.]根以及白羽扇豆(Lupinus albus)、黄花苜蓿(Medicago falcata L.)中的乙烯含量则会增加[24].在拟南芥中,缺磷能增强AtACS2、AtACS4和AtACS6的表达[25],并使编码乙烯响应转录因子ERF1和ERF2等基因的表达上调[26].研究表明,乙烯响应基因受缺磷的诱导表达在很大程度上依赖于缺磷响应核心调控因子PHR1(phosphate starvation response 1)及其同源蛋白AtPHL1[27].在phr1 phl1双突变体中,至少有8个AP2/ERF基因的表达相比于野生型下调[28].此外,缺磷也会改变植物对乙烯的敏感性.缺磷可增强玉米幼苗不定根[29]、菜豆(Phaseolus vulgaris Linn.)基根[30]和白三叶(Trifolium repens L.)侧根[31]对乙烯的敏感性,推测可能是因为缺磷改变了乙烯信号转导相关基因的表达. ...
Ethylene and plant responses to phosphate deficiency
1
2015
... 磷是植物正常生长发育和产量形成所需的大量元素之一,主要以无机磷酸根(Pi,H2PO4-)的形式被植物吸收.缺磷会改变植物乙烯的生物合成和信号转导.但缺磷对乙烯合成的影响因作物种类不同而异,如,玉米(Zea mays)[22]和番茄[23]中的乙烯含量降低,而普通大豆[Glycine max (Linn.) Merr.]根以及白羽扇豆(Lupinus albus)、黄花苜蓿(Medicago falcata L.)中的乙烯含量则会增加[24].在拟南芥中,缺磷能增强AtACS2、AtACS4和AtACS6的表达[25],并使编码乙烯响应转录因子ERF1和ERF2等基因的表达上调[26].研究表明,乙烯响应基因受缺磷的诱导表达在很大程度上依赖于缺磷响应核心调控因子PHR1(phosphate starvation response 1)及其同源蛋白AtPHL1[27].在phr1 phl1双突变体中,至少有8个AP2/ERF基因的表达相比于野生型下调[28].此外,缺磷也会改变植物对乙烯的敏感性.缺磷可增强玉米幼苗不定根[29]、菜豆(Phaseolus vulgaris Linn.)基根[30]和白三叶(Trifolium repens L.)侧根[31]对乙烯的敏感性,推测可能是因为缺磷改变了乙烯信号转导相关基因的表达. ...
A central regulatory system largely controls transcriptional activation and repression responses to phosphate starvation in Arabidopsis
1
2010
... 磷是植物正常生长发育和产量形成所需的大量元素之一,主要以无机磷酸根(Pi,H2PO4-)的形式被植物吸收.缺磷会改变植物乙烯的生物合成和信号转导.但缺磷对乙烯合成的影响因作物种类不同而异,如,玉米(Zea mays)[22]和番茄[23]中的乙烯含量降低,而普通大豆[Glycine max (Linn.) Merr.]根以及白羽扇豆(Lupinus albus)、黄花苜蓿(Medicago falcata L.)中的乙烯含量则会增加[24].在拟南芥中,缺磷能增强AtACS2、AtACS4和AtACS6的表达[25],并使编码乙烯响应转录因子ERF1和ERF2等基因的表达上调[26].研究表明,乙烯响应基因受缺磷的诱导表达在很大程度上依赖于缺磷响应核心调控因子PHR1(phosphate starvation response 1)及其同源蛋白AtPHL1[27].在phr1 phl1双突变体中,至少有8个AP2/ERF基因的表达相比于野生型下调[28].此外,缺磷也会改变植物对乙烯的敏感性.缺磷可增强玉米幼苗不定根[29]、菜豆(Phaseolus vulgaris Linn.)基根[30]和白三叶(Trifolium repens L.)侧根[31]对乙烯的敏感性,推测可能是因为缺磷改变了乙烯信号转导相关基因的表达. ...
Enhanced sensitivity to ethylene in nitrogen- or phosphate-starved roots of Zea mays L. during aerenchyma formation
2
1992
... 磷是植物正常生长发育和产量形成所需的大量元素之一,主要以无机磷酸根(Pi,H2PO4-)的形式被植物吸收.缺磷会改变植物乙烯的生物合成和信号转导.但缺磷对乙烯合成的影响因作物种类不同而异,如,玉米(Zea mays)[22]和番茄[23]中的乙烯含量降低,而普通大豆[Glycine max (Linn.) Merr.]根以及白羽扇豆(Lupinus albus)、黄花苜蓿(Medicago falcata L.)中的乙烯含量则会增加[24].在拟南芥中,缺磷能增强AtACS2、AtACS4和AtACS6的表达[25],并使编码乙烯响应转录因子ERF1和ERF2等基因的表达上调[26].研究表明,乙烯响应基因受缺磷的诱导表达在很大程度上依赖于缺磷响应核心调控因子PHR1(phosphate starvation response 1)及其同源蛋白AtPHL1[27].在phr1 phl1双突变体中,至少有8个AP2/ERF基因的表达相比于野生型下调[28].此外,缺磷也会改变植物对乙烯的敏感性.缺磷可增强玉米幼苗不定根[29]、菜豆(Phaseolus vulgaris Linn.)基根[30]和白三叶(Trifolium repens L.)侧根[31]对乙烯的敏感性,推测可能是因为缺磷改变了乙烯信号转导相关基因的表达. ...
... 硝态氮(NO3--N)和铵态氮(NH4+-N)是植物吸收的2种主要的无机氮源.除了作为营养物质,NO3--N和NH4+-N还能作为信号物质影响植物的生长发育[51].缺氮导致植物老叶变黄和生长发育迟缓[52].此外,缺氮会减少玉米根系乙烯的产生[22],但会增加根对乙烯的敏感性并诱导通气组织的形成[29],说明乙烯与氮素营养密切相关. ...
Ethylene modulates genetic, positional, and nutritional regulation of root plagiogravitropism
1
2007
... 磷是植物正常生长发育和产量形成所需的大量元素之一,主要以无机磷酸根(Pi,H2PO4-)的形式被植物吸收.缺磷会改变植物乙烯的生物合成和信号转导.但缺磷对乙烯合成的影响因作物种类不同而异,如,玉米(Zea mays)[22]和番茄[23]中的乙烯含量降低,而普通大豆[Glycine max (Linn.) Merr.]根以及白羽扇豆(Lupinus albus)、黄花苜蓿(Medicago falcata L.)中的乙烯含量则会增加[24].在拟南芥中,缺磷能增强AtACS2、AtACS4和AtACS6的表达[25],并使编码乙烯响应转录因子ERF1和ERF2等基因的表达上调[26].研究表明,乙烯响应基因受缺磷的诱导表达在很大程度上依赖于缺磷响应核心调控因子PHR1(phosphate starvation response 1)及其同源蛋白AtPHL1[27].在phr1 phl1双突变体中,至少有8个AP2/ERF基因的表达相比于野生型下调[28].此外,缺磷也会改变植物对乙烯的敏感性.缺磷可增强玉米幼苗不定根[29]、菜豆(Phaseolus vulgaris Linn.)基根[30]和白三叶(Trifolium repens L.)侧根[31]对乙烯的敏感性,推测可能是因为缺磷改变了乙烯信号转导相关基因的表达. ...
Regulation of root growth by auxin and ethylene is influenced by phosphate supply in white clover (Trifolium repens L.)
1
2012
... 磷是植物正常生长发育和产量形成所需的大量元素之一,主要以无机磷酸根(Pi,H2PO4-)的形式被植物吸收.缺磷会改变植物乙烯的生物合成和信号转导.但缺磷对乙烯合成的影响因作物种类不同而异,如,玉米(Zea mays)[22]和番茄[23]中的乙烯含量降低,而普通大豆[Glycine max (Linn.) Merr.]根以及白羽扇豆(Lupinus albus)、黄花苜蓿(Medicago falcata L.)中的乙烯含量则会增加[24].在拟南芥中,缺磷能增强AtACS2、AtACS4和AtACS6的表达[25],并使编码乙烯响应转录因子ERF1和ERF2等基因的表达上调[26].研究表明,乙烯响应基因受缺磷的诱导表达在很大程度上依赖于缺磷响应核心调控因子PHR1(phosphate starvation response 1)及其同源蛋白AtPHL1[27].在phr1 phl1双突变体中,至少有8个AP2/ERF基因的表达相比于野生型下调[28].此外,缺磷也会改变植物对乙烯的敏感性.缺磷可增强玉米幼苗不定根[29]、菜豆(Phaseolus vulgaris Linn.)基根[30]和白三叶(Trifolium repens L.)侧根[31]对乙烯的敏感性,推测可能是因为缺磷改变了乙烯信号转导相关基因的表达. ...
Light and ethylene coordinately regulate the phosphate starvation response through transcriptional regulation of PHOSPHATE STARVATION RESPONSE1
1
2017
... 在分子生理水平上,乙烯通过调节磷吸收、活化和再利用相关基因来调控植物缺磷响应.磷转运蛋白基因PT1(PHT1;1)和PT2(PHT1;4)的表达水平在拟南芥乙烯不敏感突变体etr1和ein2-5中显著降低,而在拟南芥组成型乙烯反应突变体hps2(hypersensitive to Pi starvation 2,ctr1同源物)中显著增加[25].已有研究表明,ACC可以诱导缺磷响应核心调控因子PHR1的表达,后者调控一系列磷吸收、转运和饥饿应答相关基因的表达[32].EIN3/EIL1通过调控转录因子WRKY75激活PHT1;5的表达[33].在其他植物中也有类似的发现,例如在苜蓿中,ACC在磷充足的条件下诱导磷转运蛋白基因MfPT1和MfPT5的表达,而乙烯生物合成抑制剂氨基乙氧基乙烯基甘氨酸(aminoethoxyvinylglycine, AVG)和Co2+均阻止了这些基因的低磷诱导表达[34].在低磷胁迫下,植物通过分泌酸性磷酸酶(acid phosphatase, APase)来活化土壤中的磷,以提高对有机磷的利用率.研究发现,拟南芥hps2突变体根表面的APase活性明显增加,而其ein2突变体的APase活性则显著降低,表明乙烯是APase活性的正调控因子[25].此外,花青素累积是缺磷的标志性症状,在低磷条件下,拟南芥hps2、hps3和hps4突变体中花青素的积累量低于野生型[25,35],但在ein2突变体中花青素含量则明显增加,说明乙烯是磷饥饿诱导花青素积累的负调控因子[25].hps2、hps3等突变体中花青素的减少可能是由于激活的乙烯信号通路缓解了低磷胁迫[25].在体内磷再利用方面,乙烯可促进拟南芥细胞壁磷的再利用,NAC转录因子ANAC044作为转录抑制因子,通过下调ACS6和ACO1的表达来抑制缺磷诱导的乙烯生物合成,从而抑制细胞壁磷的再利用[36]. ...
A tripartite amplification loop involving the transcription factor WRKY75, salicylic acid, and reactive oxygen species accelerates leaf senescence
1
2017
... 在分子生理水平上,乙烯通过调节磷吸收、活化和再利用相关基因来调控植物缺磷响应.磷转运蛋白基因PT1(PHT1;1)和PT2(PHT1;4)的表达水平在拟南芥乙烯不敏感突变体etr1和ein2-5中显著降低,而在拟南芥组成型乙烯反应突变体hps2(hypersensitive to Pi starvation 2,ctr1同源物)中显著增加[25].已有研究表明,ACC可以诱导缺磷响应核心调控因子PHR1的表达,后者调控一系列磷吸收、转运和饥饿应答相关基因的表达[32].EIN3/EIL1通过调控转录因子WRKY75激活PHT1;5的表达[33].在其他植物中也有类似的发现,例如在苜蓿中,ACC在磷充足的条件下诱导磷转运蛋白基因MfPT1和MfPT5的表达,而乙烯生物合成抑制剂氨基乙氧基乙烯基甘氨酸(aminoethoxyvinylglycine, AVG)和Co2+均阻止了这些基因的低磷诱导表达[34].在低磷胁迫下,植物通过分泌酸性磷酸酶(acid phosphatase, APase)来活化土壤中的磷,以提高对有机磷的利用率.研究发现,拟南芥hps2突变体根表面的APase活性明显增加,而其ein2突变体的APase活性则显著降低,表明乙烯是APase活性的正调控因子[25].此外,花青素累积是缺磷的标志性症状,在低磷条件下,拟南芥hps2、hps3和hps4突变体中花青素的积累量低于野生型[25,35],但在ein2突变体中花青素含量则明显增加,说明乙烯是磷饥饿诱导花青素积累的负调控因子[25].hps2、hps3等突变体中花青素的减少可能是由于激活的乙烯信号通路缓解了低磷胁迫[25].在体内磷再利用方面,乙烯可促进拟南芥细胞壁磷的再利用,NAC转录因子ANAC044作为转录抑制因子,通过下调ACS6和ACO1的表达来抑制缺磷诱导的乙烯生物合成,从而抑制细胞壁磷的再利用[36]. ...
Stimulation of root acid phosphatase by phosphorus deficiency is regulated by ethylene in Medicago falcata
1
2011
... 在分子生理水平上,乙烯通过调节磷吸收、活化和再利用相关基因来调控植物缺磷响应.磷转运蛋白基因PT1(PHT1;1)和PT2(PHT1;4)的表达水平在拟南芥乙烯不敏感突变体etr1和ein2-5中显著降低,而在拟南芥组成型乙烯反应突变体hps2(hypersensitive to Pi starvation 2,ctr1同源物)中显著增加[25].已有研究表明,ACC可以诱导缺磷响应核心调控因子PHR1的表达,后者调控一系列磷吸收、转运和饥饿应答相关基因的表达[32].EIN3/EIL1通过调控转录因子WRKY75激活PHT1;5的表达[33].在其他植物中也有类似的发现,例如在苜蓿中,ACC在磷充足的条件下诱导磷转运蛋白基因MfPT1和MfPT5的表达,而乙烯生物合成抑制剂氨基乙氧基乙烯基甘氨酸(aminoethoxyvinylglycine, AVG)和Co2+均阻止了这些基因的低磷诱导表达[34].在低磷胁迫下,植物通过分泌酸性磷酸酶(acid phosphatase, APase)来活化土壤中的磷,以提高对有机磷的利用率.研究发现,拟南芥hps2突变体根表面的APase活性明显增加,而其ein2突变体的APase活性则显著降低,表明乙烯是APase活性的正调控因子[25].此外,花青素累积是缺磷的标志性症状,在低磷条件下,拟南芥hps2、hps3和hps4突变体中花青素的积累量低于野生型[25,35],但在ein2突变体中花青素含量则明显增加,说明乙烯是磷饥饿诱导花青素积累的负调控因子[25].hps2、hps3等突变体中花青素的减少可能是由于激活的乙烯信号通路缓解了低磷胁迫[25].在体内磷再利用方面,乙烯可促进拟南芥细胞壁磷的再利用,NAC转录因子ANAC044作为转录抑制因子,通过下调ACS6和ACO1的表达来抑制缺磷诱导的乙烯生物合成,从而抑制细胞壁磷的再利用[36]. ...
The Arabidopsis gene HYPERSENSITIVE TO PHOSPHATE STARVATION 3 encodes ETHYLENE OVER PRODUCTION 1
1
2012
... 在分子生理水平上,乙烯通过调节磷吸收、活化和再利用相关基因来调控植物缺磷响应.磷转运蛋白基因PT1(PHT1;1)和PT2(PHT1;4)的表达水平在拟南芥乙烯不敏感突变体etr1和ein2-5中显著降低,而在拟南芥组成型乙烯反应突变体hps2(hypersensitive to Pi starvation 2,ctr1同源物)中显著增加[25].已有研究表明,ACC可以诱导缺磷响应核心调控因子PHR1的表达,后者调控一系列磷吸收、转运和饥饿应答相关基因的表达[32].EIN3/EIL1通过调控转录因子WRKY75激活PHT1;5的表达[33].在其他植物中也有类似的发现,例如在苜蓿中,ACC在磷充足的条件下诱导磷转运蛋白基因MfPT1和MfPT5的表达,而乙烯生物合成抑制剂氨基乙氧基乙烯基甘氨酸(aminoethoxyvinylglycine, AVG)和Co2+均阻止了这些基因的低磷诱导表达[34].在低磷胁迫下,植物通过分泌酸性磷酸酶(acid phosphatase, APase)来活化土壤中的磷,以提高对有机磷的利用率.研究发现,拟南芥hps2突变体根表面的APase活性明显增加,而其ein2突变体的APase活性则显著降低,表明乙烯是APase活性的正调控因子[25].此外,花青素累积是缺磷的标志性症状,在低磷条件下,拟南芥hps2、hps3和hps4突变体中花青素的积累量低于野生型[25,35],但在ein2突变体中花青素含量则明显增加,说明乙烯是磷饥饿诱导花青素积累的负调控因子[25].hps2、hps3等突变体中花青素的减少可能是由于激活的乙烯信号通路缓解了低磷胁迫[25].在体内磷再利用方面,乙烯可促进拟南芥细胞壁磷的再利用,NAC转录因子ANAC044作为转录抑制因子,通过下调ACS6和ACO1的表达来抑制缺磷诱导的乙烯生物合成,从而抑制细胞壁磷的再利用[36]. ...
ANAC044 is associated with P reutilization in P deficient Arabidopsis thaliana root cell wall in an ethylene dependent manner
1
2021
... 在分子生理水平上,乙烯通过调节磷吸收、活化和再利用相关基因来调控植物缺磷响应.磷转运蛋白基因PT1(PHT1;1)和PT2(PHT1;4)的表达水平在拟南芥乙烯不敏感突变体etr1和ein2-5中显著降低,而在拟南芥组成型乙烯反应突变体hps2(hypersensitive to Pi starvation 2,ctr1同源物)中显著增加[25].已有研究表明,ACC可以诱导缺磷响应核心调控因子PHR1的表达,后者调控一系列磷吸收、转运和饥饿应答相关基因的表达[32].EIN3/EIL1通过调控转录因子WRKY75激活PHT1;5的表达[33].在其他植物中也有类似的发现,例如在苜蓿中,ACC在磷充足的条件下诱导磷转运蛋白基因MfPT1和MfPT5的表达,而乙烯生物合成抑制剂氨基乙氧基乙烯基甘氨酸(aminoethoxyvinylglycine, AVG)和Co2+均阻止了这些基因的低磷诱导表达[34].在低磷胁迫下,植物通过分泌酸性磷酸酶(acid phosphatase, APase)来活化土壤中的磷,以提高对有机磷的利用率.研究发现,拟南芥hps2突变体根表面的APase活性明显增加,而其ein2突变体的APase活性则显著降低,表明乙烯是APase活性的正调控因子[25].此外,花青素累积是缺磷的标志性症状,在低磷条件下,拟南芥hps2、hps3和hps4突变体中花青素的积累量低于野生型[25,35],但在ein2突变体中花青素含量则明显增加,说明乙烯是磷饥饿诱导花青素积累的负调控因子[25].hps2、hps3等突变体中花青素的减少可能是由于激活的乙烯信号通路缓解了低磷胁迫[25].在体内磷再利用方面,乙烯可促进拟南芥细胞壁磷的再利用,NAC转录因子ANAC044作为转录抑制因子,通过下调ACS6和ACO1的表达来抑制缺磷诱导的乙烯生物合成,从而抑制细胞壁磷的再利用[36]. ...
Nitric oxide acts upstream of ethylene in cell wall phosphorus reutilization in phosphorus-deficient rice
1
2017
... 在形态发育水平上,植物可以通过重塑根系构型应对缺磷胁迫,包括抑制主根生长、刺激侧根发育与伸长、增加根毛数量、促进排根形成等[37].施用Ag+或AVG可以抑制低磷诱导的主根分生组织衰竭,表明乙烯参与了缺磷导致的主根生长抑制过程[38].研究发现,GmETO1(ethylene-overproduction protein 1 in G. max)通过调节乙烯合成和下游相关基因的表达,影响大豆毛状根的增殖和伸长,提高磷的吸收利用率,进而增强大豆对低磷胁迫的耐受性[39].缺磷诱导EIN3蛋白积累,而EIN3一方面直接调控RSL4(ROOT HAIR DEFECTIVE 6-LIKE 4)的表达[19],另一方面通过结合到一组调节细胞壁修饰的基因,如RHS19、EXT14的启动子上,激活它们的转录,进而促进根毛的发育[40]. ...
Global expression pattern comparison between low phosphorus insensitive 4 and WT Arabidopsis reveals an important role of reactive oxygen species and jasmonic acid in the root tip response to phosphate starvation
1
2011
... 在形态发育水平上,植物可以通过重塑根系构型应对缺磷胁迫,包括抑制主根生长、刺激侧根发育与伸长、增加根毛数量、促进排根形成等[37].施用Ag+或AVG可以抑制低磷诱导的主根分生组织衰竭,表明乙烯参与了缺磷导致的主根生长抑制过程[38].研究发现,GmETO1(ethylene-overproduction protein 1 in G. max)通过调节乙烯合成和下游相关基因的表达,影响大豆毛状根的增殖和伸长,提高磷的吸收利用率,进而增强大豆对低磷胁迫的耐受性[39].缺磷诱导EIN3蛋白积累,而EIN3一方面直接调控RSL4(ROOT HAIR DEFECTIVE 6-LIKE 4)的表达[19],另一方面通过结合到一组调节细胞壁修饰的基因,如RHS19、EXT14的启动子上,激活它们的转录,进而促进根毛的发育[40]. ...
Up-regulating GmETO1 improves phosphorus uptake and use efficiency by promoting root growth in soybean
1
2020
... 在形态发育水平上,植物可以通过重塑根系构型应对缺磷胁迫,包括抑制主根生长、刺激侧根发育与伸长、增加根毛数量、促进排根形成等[37].施用Ag+或AVG可以抑制低磷诱导的主根分生组织衰竭,表明乙烯参与了缺磷导致的主根生长抑制过程[38].研究发现,GmETO1(ethylene-overproduction protein 1 in G. max)通过调节乙烯合成和下游相关基因的表达,影响大豆毛状根的增殖和伸长,提高磷的吸收利用率,进而增强大豆对低磷胁迫的耐受性[39].缺磷诱导EIN3蛋白积累,而EIN3一方面直接调控RSL4(ROOT HAIR DEFECTIVE 6-LIKE 4)的表达[19],另一方面通过结合到一组调节细胞壁修饰的基因,如RHS19、EXT14的启动子上,激活它们的转录,进而促进根毛的发育[40]. ...
The molecular mechanism of ethylene-mediated root hair development induced by phosphate starvation
1
2016
... 在形态发育水平上,植物可以通过重塑根系构型应对缺磷胁迫,包括抑制主根生长、刺激侧根发育与伸长、增加根毛数量、促进排根形成等[37].施用Ag+或AVG可以抑制低磷诱导的主根分生组织衰竭,表明乙烯参与了缺磷导致的主根生长抑制过程[38].研究发现,GmETO1(ethylene-overproduction protein 1 in G. max)通过调节乙烯合成和下游相关基因的表达,影响大豆毛状根的增殖和伸长,提高磷的吸收利用率,进而增强大豆对低磷胁迫的耐受性[39].缺磷诱导EIN3蛋白积累,而EIN3一方面直接调控RSL4(ROOT HAIR DEFECTIVE 6-LIKE 4)的表达[19],另一方面通过结合到一组调节细胞壁修饰的基因,如RHS19、EXT14的启动子上,激活它们的转录,进而促进根毛的发育[40]. ...
Potassium: a key modulator for cell homeostasis
1
2020
... 钾是植物必需的大量营养元素,是植物细胞中最丰富的阳离子,缺钾严重影响植物生长和产量[41].乙烯信号通路是植物对低钾反应的关键组成部分,刺激活性氧(reactive oxygen species, ROS)产生,诱导根系形态改变以及高亲和力转运蛋白基因的表达[42].在缺K条件下,拟南芥幼苗释放的乙烯量明显增加,这与缺钾诱导ACS2和ACS6基因表达上调直接相关[42].此外,西瓜[Citrullus lanatus (Thunb.) Matsum. et Nakai]中的ERF1和EBF1也受缺钾诱导表达[43].研究发现,乙烯是缺钾/低钾诱导ROS产生的重要前提,而ROS可以增强高亲和K+转运蛋白基因HAK5的表达,而在乙烯信号缺失突变体中,低钾诱导的ROS含量和HAK5表达量则会显著降低[42,44],说明乙烯在缺钾/低钾条件下增强植物对钾的吸收利用发挥着重要的正向调控作用. ...
Ethylene mediates response and tolerance to potassium deprivation in Arabidopsis
5
2009
... 钾是植物必需的大量营养元素,是植物细胞中最丰富的阳离子,缺钾严重影响植物生长和产量[41].乙烯信号通路是植物对低钾反应的关键组成部分,刺激活性氧(reactive oxygen species, ROS)产生,诱导根系形态改变以及高亲和力转运蛋白基因的表达[42].在缺K条件下,拟南芥幼苗释放的乙烯量明显增加,这与缺钾诱导ACS2和ACS6基因表达上调直接相关[42].此外,西瓜[Citrullus lanatus (Thunb.) Matsum. et Nakai]中的ERF1和EBF1也受缺钾诱导表达[43].研究发现,乙烯是缺钾/低钾诱导ROS产生的重要前提,而ROS可以增强高亲和K+转运蛋白基因HAK5的表达,而在乙烯信号缺失突变体中,低钾诱导的ROS含量和HAK5表达量则会显著降低[42,44],说明乙烯在缺钾/低钾条件下增强植物对钾的吸收利用发挥着重要的正向调控作用. ...
... [42].此外,西瓜[Citrullus lanatus (Thunb.) Matsum. et Nakai]中的ERF1和EBF1也受缺钾诱导表达[43].研究发现,乙烯是缺钾/低钾诱导ROS产生的重要前提,而ROS可以增强高亲和K+转运蛋白基因HAK5的表达,而在乙烯信号缺失突变体中,低钾诱导的ROS含量和HAK5表达量则会显著降低[42,44],说明乙烯在缺钾/低钾条件下增强植物对钾的吸收利用发挥着重要的正向调控作用. ...
... [42,44],说明乙烯在缺钾/低钾条件下增强植物对钾的吸收利用发挥着重要的正向调控作用. ...
... 在形态发育水平上,乙烯参与调控缺钾条件下植物的根系发育.缺钾抑制主根生长,促进根毛伸长[42].研究发现,锌指蛋白5(zinc finger protein 5, ZFP5)转录因子通过激活拟南芥中EIN2的表达,正向调控根毛的伸长以响应磷和钾的缺乏[45].但在低钾条件下,拟南芥突变体ein2和etr1的主根生长并没有被抑制,仍能刺激ROS产生和根毛生长,表明其他信号通路或不依赖EIN2的乙烯通路也参与了低钾条件下的根系发育[42]. ...
... [42]. ...
Compressibility and hydraulic conductivity of clayey soil mixed with calcium bentonite for slurry wall backfill: initial assessment
1
2014
... 钾是植物必需的大量营养元素,是植物细胞中最丰富的阳离子,缺钾严重影响植物生长和产量[41].乙烯信号通路是植物对低钾反应的关键组成部分,刺激活性氧(reactive oxygen species, ROS)产生,诱导根系形态改变以及高亲和力转运蛋白基因的表达[42].在缺K条件下,拟南芥幼苗释放的乙烯量明显增加,这与缺钾诱导ACS2和ACS6基因表达上调直接相关[42].此外,西瓜[Citrullus lanatus (Thunb.) Matsum. et Nakai]中的ERF1和EBF1也受缺钾诱导表达[43].研究发现,乙烯是缺钾/低钾诱导ROS产生的重要前提,而ROS可以增强高亲和K+转运蛋白基因HAK5的表达,而在乙烯信号缺失突变体中,低钾诱导的ROS含量和HAK5表达量则会显著降低[42,44],说明乙烯在缺钾/低钾条件下增强植物对钾的吸收利用发挥着重要的正向调控作用. ...
Molecular mechanisms involved in plant adaptation to low K+ availability
1
2014
... 钾是植物必需的大量营养元素,是植物细胞中最丰富的阳离子,缺钾严重影响植物生长和产量[41].乙烯信号通路是植物对低钾反应的关键组成部分,刺激活性氧(reactive oxygen species, ROS)产生,诱导根系形态改变以及高亲和力转运蛋白基因的表达[42].在缺K条件下,拟南芥幼苗释放的乙烯量明显增加,这与缺钾诱导ACS2和ACS6基因表达上调直接相关[42].此外,西瓜[Citrullus lanatus (Thunb.) Matsum. et Nakai]中的ERF1和EBF1也受缺钾诱导表达[43].研究发现,乙烯是缺钾/低钾诱导ROS产生的重要前提,而ROS可以增强高亲和K+转运蛋白基因HAK5的表达,而在乙烯信号缺失突变体中,低钾诱导的ROS含量和HAK5表达量则会显著降低[42,44],说明乙烯在缺钾/低钾条件下增强植物对钾的吸收利用发挥着重要的正向调控作用. ...
Zinc finger protein 5 (ZFP5) associates with ethylene signaling to regulate the phosphate and potassium deficiency-induced root hair development in Arabidopsis
1
2020
... 在形态发育水平上,乙烯参与调控缺钾条件下植物的根系发育.缺钾抑制主根生长,促进根毛伸长[42].研究发现,锌指蛋白5(zinc finger protein 5, ZFP5)转录因子通过激活拟南芥中EIN2的表达,正向调控根毛的伸长以响应磷和钾的缺乏[45].但在低钾条件下,拟南芥突变体ein2和etr1的主根生长并没有被抑制,仍能刺激ROS产生和根毛生长,表明其他信号通路或不依赖EIN2的乙烯通路也参与了低钾条件下的根系发育[42]. ...
Links between ethylene and sulfur nutrition: a regulatory interplay or just metabolite association?
2
2015
... 硫是植物必需的大量元素之一,植物根系以硫酸盐的形式吸收,然后将其还原并用于一系列初级和次级代谢产物的合成[46].硫代谢物SAM可用于合成乙烯前体ACC[46].缺硫植物的新叶发黄,当给缺硫的油菜供应乙烯后,其叶绿素含量增加,表明乙烯可以缓解缺硫症状[47]. ...
... [46].缺硫植物的新叶发黄,当给缺硫的油菜供应乙烯后,其叶绿素含量增加,表明乙烯可以缓解缺硫症状[47]. ...
Ethylene regulates sulfur acquisition by regulating the expression of sulfate transporter genes in oilseed rape
2
2021
... 硫是植物必需的大量元素之一,植物根系以硫酸盐的形式吸收,然后将其还原并用于一系列初级和次级代谢产物的合成[46].硫代谢物SAM可用于合成乙烯前体ACC[46].缺硫植物的新叶发黄,当给缺硫的油菜供应乙烯后,其叶绿素含量增加,表明乙烯可以缓解缺硫症状[47]. ...
... SLIM1(sulfur limitation 1)是在缺硫条件下调节硫酸盐吸收以及含硫次生代谢物循环的关键转录因子,而SLIM1是6个EIL成员之一,也称为EIL3[48].研究发现,EIN3与SLIM1形成的异源二聚体破坏了SLIM1的DNA结合能力,进而干扰拟南芥的缺硫响应[49].除了SLIM1,其他EIL成员也参与缺硫响应,如:在正常生长条件下,EIL1或EIL3的突变影响了含硫次生代谢物的积累,且这一现象在缺硫条件下更加明显[50];此外,EIL1和EIL3还参与调控植物体内硫酸盐的运输[50].但比较EIL1和SLIM1/EIL3功能缺失后的差异表达基因却发现,在eil3突变体中有598个差异表达基因,而在eil1中仅有72个,可以推测SLIM1/EIL3在硫响应调控中更为重要,而EIL1可能起辅助作用[50].在油菜中,参与硫转运和调控的基因(如SULTRs、ATP sulfurylase、OASTL等)在缺硫条件下也受到乙烯的调控[47]. ...
Arabidopsis SLIM1 is a central transcriptional regulator of plant sulfur response and metabolism
1
2006
... SLIM1(sulfur limitation 1)是在缺硫条件下调节硫酸盐吸收以及含硫次生代谢物循环的关键转录因子,而SLIM1是6个EIL成员之一,也称为EIL3[48].研究发现,EIN3与SLIM1形成的异源二聚体破坏了SLIM1的DNA结合能力,进而干扰拟南芥的缺硫响应[49].除了SLIM1,其他EIL成员也参与缺硫响应,如:在正常生长条件下,EIL1或EIL3的突变影响了含硫次生代谢物的积累,且这一现象在缺硫条件下更加明显[50];此外,EIL1和EIL3还参与调控植物体内硫酸盐的运输[50].但比较EIL1和SLIM1/EIL3功能缺失后的差异表达基因却发现,在eil3突变体中有598个差异表达基因,而在eil1中仅有72个,可以推测SLIM1/EIL3在硫响应调控中更为重要,而EIL1可能起辅助作用[50].在油菜中,参与硫转运和调控的基因(如SULTRs、ATP sulfurylase、OASTL等)在缺硫条件下也受到乙烯的调控[47]. ...
EIN3 interferes with the sulfur deficiency signaling in Arabidopsis thaliana through direct interaction with the SLIM1 transcription factor
1
2016
... SLIM1(sulfur limitation 1)是在缺硫条件下调节硫酸盐吸收以及含硫次生代谢物循环的关键转录因子,而SLIM1是6个EIL成员之一,也称为EIL3[48].研究发现,EIN3与SLIM1形成的异源二聚体破坏了SLIM1的DNA结合能力,进而干扰拟南芥的缺硫响应[49].除了SLIM1,其他EIL成员也参与缺硫响应,如:在正常生长条件下,EIL1或EIL3的突变影响了含硫次生代谢物的积累,且这一现象在缺硫条件下更加明显[50];此外,EIL1和EIL3还参与调控植物体内硫酸盐的运输[50].但比较EIL1和SLIM1/EIL3功能缺失后的差异表达基因却发现,在eil3突变体中有598个差异表达基因,而在eil1中仅有72个,可以推测SLIM1/EIL3在硫响应调控中更为重要,而EIL1可能起辅助作用[50].在油菜中,参与硫转运和调控的基因(如SULTRs、ATP sulfurylase、OASTL等)在缺硫条件下也受到乙烯的调控[47]. ...
The transcription factor EIL1 participates in the regulation of sulfur-deficiency response
3
2020
... SLIM1(sulfur limitation 1)是在缺硫条件下调节硫酸盐吸收以及含硫次生代谢物循环的关键转录因子,而SLIM1是6个EIL成员之一,也称为EIL3[48].研究发现,EIN3与SLIM1形成的异源二聚体破坏了SLIM1的DNA结合能力,进而干扰拟南芥的缺硫响应[49].除了SLIM1,其他EIL成员也参与缺硫响应,如:在正常生长条件下,EIL1或EIL3的突变影响了含硫次生代谢物的积累,且这一现象在缺硫条件下更加明显[50];此外,EIL1和EIL3还参与调控植物体内硫酸盐的运输[50].但比较EIL1和SLIM1/EIL3功能缺失后的差异表达基因却发现,在eil3突变体中有598个差异表达基因,而在eil1中仅有72个,可以推测SLIM1/EIL3在硫响应调控中更为重要,而EIL1可能起辅助作用[50].在油菜中,参与硫转运和调控的基因(如SULTRs、ATP sulfurylase、OASTL等)在缺硫条件下也受到乙烯的调控[47]. ...
... [50].但比较EIL1和SLIM1/EIL3功能缺失后的差异表达基因却发现,在eil3突变体中有598个差异表达基因,而在eil1中仅有72个,可以推测SLIM1/EIL3在硫响应调控中更为重要,而EIL1可能起辅助作用[50].在油菜中,参与硫转运和调控的基因(如SULTRs、ATP sulfurylase、OASTL等)在缺硫条件下也受到乙烯的调控[47]. ...
... [50].在油菜中,参与硫转运和调控的基因(如SULTRs、ATP sulfurylase、OASTL等)在缺硫条件下也受到乙烯的调控[47]. ...
Nitrogen form-mediated ethylene signal regulates root-to-shoot K+ translocation via NRT1.5
3
2021
... 硝态氮(NO3--N)和铵态氮(NH4+-N)是植物吸收的2种主要的无机氮源.除了作为营养物质,NO3--N和NH4+-N还能作为信号物质影响植物的生长发育[51].缺氮导致植物老叶变黄和生长发育迟缓[52].此外,缺氮会减少玉米根系乙烯的产生[22],但会增加根对乙烯的敏感性并诱导通气组织的形成[29],说明乙烯与氮素营养密切相关. ...
... 研究发现,硝酸盐缺乏能诱导植物侧根伸长[53],同时,通过诱导EIN3和EIL1的表达来触发乙烯生物合成并激活乙烯信号[54].在低硝酸盐条件下,NRT2.1(NITRATE TRANSPORTER 2.1)表达上调并可能以NRT2.1依赖的方式诱导乙烯的生物合成和信号转导,而乙烯反过来抑制NRT2.1表达,减少硝酸盐的高亲和吸收,这是一种负反馈调控,旨在平衡植物对硝酸盐的吸收[55].NH4+-N促进了乙烯的生物合成和增强了EIN3的转录活性,进而降低钾转运蛋白基因AtNRT1.5的转录水平以抑制K+从根到地上部的转运,最终造成叶片提前衰老,这也可能是NH4+-N过量供应抑制K+吸收的根本原因[51];而供应少量的NO3--N可以强烈抑制EIN3的活性,并显著增加AtNRT1.5的表达和K+从根到茎的转移,充分说明乙烯通过其核心转录因子EIN3在调控氮钾互作中发挥重要作用[51]. ...
... [51]. ...
Current understanding on ethylene signaling in plants: the influence of nutrient availability
1
2013
... 硝态氮(NO3--N)和铵态氮(NH4+-N)是植物吸收的2种主要的无机氮源.除了作为营养物质,NO3--N和NH4+-N还能作为信号物质影响植物的生长发育[51].缺氮导致植物老叶变黄和生长发育迟缓[52].此外,缺氮会减少玉米根系乙烯的产生[22],但会增加根对乙烯的敏感性并诱导通气组织的形成[29],说明乙烯与氮素营养密切相关. ...
Role of ethylene in responses of plants to nitrogen availability
1
2015
... 研究发现,硝酸盐缺乏能诱导植物侧根伸长[53],同时,通过诱导EIN3和EIL1的表达来触发乙烯生物合成并激活乙烯信号[54].在低硝酸盐条件下,NRT2.1(NITRATE TRANSPORTER 2.1)表达上调并可能以NRT2.1依赖的方式诱导乙烯的生物合成和信号转导,而乙烯反过来抑制NRT2.1表达,减少硝酸盐的高亲和吸收,这是一种负反馈调控,旨在平衡植物对硝酸盐的吸收[55].NH4+-N促进了乙烯的生物合成和增强了EIN3的转录活性,进而降低钾转运蛋白基因AtNRT1.5的转录水平以抑制K+从根到地上部的转运,最终造成叶片提前衰老,这也可能是NH4+-N过量供应抑制K+吸收的根本原因[51];而供应少量的NO3--N可以强烈抑制EIN3的活性,并显著增加AtNRT1.5的表达和K+从根到茎的转移,充分说明乙烯通过其核心转录因子EIN3在调控氮钾互作中发挥重要作用[51]. ...
Nitrate in 2020: thirty years from transport to signaling networks
1
2020
... 研究发现,硝酸盐缺乏能诱导植物侧根伸长[53],同时,通过诱导EIN3和EIL1的表达来触发乙烯生物合成并激活乙烯信号[54].在低硝酸盐条件下,NRT2.1(NITRATE TRANSPORTER 2.1)表达上调并可能以NRT2.1依赖的方式诱导乙烯的生物合成和信号转导,而乙烯反过来抑制NRT2.1表达,减少硝酸盐的高亲和吸收,这是一种负反馈调控,旨在平衡植物对硝酸盐的吸收[55].NH4+-N促进了乙烯的生物合成和增强了EIN3的转录活性,进而降低钾转运蛋白基因AtNRT1.5的转录水平以抑制K+从根到地上部的转运,最终造成叶片提前衰老,这也可能是NH4+-N过量供应抑制K+吸收的根本原因[51];而供应少量的NO3--N可以强烈抑制EIN3的活性,并显著增加AtNRT1.5的表达和K+从根到茎的转移,充分说明乙烯通过其核心转录因子EIN3在调控氮钾互作中发挥重要作用[51]. ...
The nitrate transporter NRT2.1 functions in the ethylene response to nitrate deficiency in Arabidopsis
1
2013
... 研究发现,硝酸盐缺乏能诱导植物侧根伸长[53],同时,通过诱导EIN3和EIL1的表达来触发乙烯生物合成并激活乙烯信号[54].在低硝酸盐条件下,NRT2.1(NITRATE TRANSPORTER 2.1)表达上调并可能以NRT2.1依赖的方式诱导乙烯的生物合成和信号转导,而乙烯反过来抑制NRT2.1表达,减少硝酸盐的高亲和吸收,这是一种负反馈调控,旨在平衡植物对硝酸盐的吸收[55].NH4+-N促进了乙烯的生物合成和增强了EIN3的转录活性,进而降低钾转运蛋白基因AtNRT1.5的转录水平以抑制K+从根到地上部的转运,最终造成叶片提前衰老,这也可能是NH4+-N过量供应抑制K+吸收的根本原因[51];而供应少量的NO3--N可以强烈抑制EIN3的活性,并显著增加AtNRT1.5的表达和K+从根到茎的转移,充分说明乙烯通过其核心转录因子EIN3在调控氮钾互作中发挥重要作用[51]. ...
HPS4/SABRE regulates plant responses to phosphate starvation through antagonistic interaction with ethylene signalling
1
2012
... 乙烯通过促进生长素(auxin, IAA)的生物合成和运输来调节根的发育.在缺磷条件下,拟南芥乙烯组成型响应突变体hps4的主根生长抑制更为严重.研究发现,hps4中生长素生物合成相关基因的表达量增加,其根尖的生长素合成量是野生型的2倍,用Ag+处理可抑制hps4对缺磷的过敏反应,而用AVG处理则不能,表明hps4中增强的乙烯信号促进了根尖生长素的生物合成,而生长素过量累积抑制了主根的生长[56].在低钾条件下,乙烯促进生长素转运蛋白基因PIN3(PIN-FORMED 3)和PIN7的表达,导致生长素输出量增加,从而阻止驱动侧根形成所需的生长素的局部积累,最终抑制侧根发育[57].此外,在低铁条件下,乙烯和生长素协同抑制根的伸长,并拮抗调控侧根的形成:生长素诱导侧根发育,而乙烯抑制侧根发育[57-58].缺硼(B)会导致乙烯和生长素介导的主根生长受到抑制,其机制可能是缺硼诱导ACS11基因表达,导致乙烯含量增加,而乙烯通过促进生长素的生物合成以及调控生长素的运输机制,激活了局部生长素信号转导,最终导致根细胞长度变短,主根生长受到抑制[59-60]. ...
Ethylene inhibits lateral root development, increases IAA transport and expres-sion of PIN3 and PIN7 auxin efflux carriers
2
2011
... 乙烯通过促进生长素(auxin, IAA)的生物合成和运输来调节根的发育.在缺磷条件下,拟南芥乙烯组成型响应突变体hps4的主根生长抑制更为严重.研究发现,hps4中生长素生物合成相关基因的表达量增加,其根尖的生长素合成量是野生型的2倍,用Ag+处理可抑制hps4对缺磷的过敏反应,而用AVG处理则不能,表明hps4中增强的乙烯信号促进了根尖生长素的生物合成,而生长素过量累积抑制了主根的生长[56].在低钾条件下,乙烯促进生长素转运蛋白基因PIN3(PIN-FORMED 3)和PIN7的表达,导致生长素输出量增加,从而阻止驱动侧根形成所需的生长素的局部积累,最终抑制侧根发育[57].此外,在低铁条件下,乙烯和生长素协同抑制根的伸长,并拮抗调控侧根的形成:生长素诱导侧根发育,而乙烯抑制侧根发育[57-58].缺硼(B)会导致乙烯和生长素介导的主根生长受到抑制,其机制可能是缺硼诱导ACS11基因表达,导致乙烯含量增加,而乙烯通过促进生长素的生物合成以及调控生长素的运输机制,激活了局部生长素信号转导,最终导致根细胞长度变短,主根生长受到抑制[59-60]. ...
... [57-58].缺硼(B)会导致乙烯和生长素介导的主根生长受到抑制,其机制可能是缺硼诱导ACS11基因表达,导致乙烯含量增加,而乙烯通过促进生长素的生物合成以及调控生长素的运输机制,激活了局部生长素信号转导,最终导致根细胞长度变短,主根生长受到抑制[59-60]. ...
The role of nutrient availability in regulating root architecture
1
2003
... 乙烯通过促进生长素(auxin, IAA)的生物合成和运输来调节根的发育.在缺磷条件下,拟南芥乙烯组成型响应突变体hps4的主根生长抑制更为严重.研究发现,hps4中生长素生物合成相关基因的表达量增加,其根尖的生长素合成量是野生型的2倍,用Ag+处理可抑制hps4对缺磷的过敏反应,而用AVG处理则不能,表明hps4中增强的乙烯信号促进了根尖生长素的生物合成,而生长素过量累积抑制了主根的生长[56].在低钾条件下,乙烯促进生长素转运蛋白基因PIN3(PIN-FORMED 3)和PIN7的表达,导致生长素输出量增加,从而阻止驱动侧根形成所需的生长素的局部积累,最终抑制侧根发育[57].此外,在低铁条件下,乙烯和生长素协同抑制根的伸长,并拮抗调控侧根的形成:生长素诱导侧根发育,而乙烯抑制侧根发育[57-58].缺硼(B)会导致乙烯和生长素介导的主根生长受到抑制,其机制可能是缺硼诱导ACS11基因表达,导致乙烯含量增加,而乙烯通过促进生长素的生物合成以及调控生长素的运输机制,激活了局部生长素信号转导,最终导致根细胞长度变短,主根生长受到抑制[59-60]. ...
Auxin and ethylene: collaborators or competitors?
1
2012
... 乙烯通过促进生长素(auxin, IAA)的生物合成和运输来调节根的发育.在缺磷条件下,拟南芥乙烯组成型响应突变体hps4的主根生长抑制更为严重.研究发现,hps4中生长素生物合成相关基因的表达量增加,其根尖的生长素合成量是野生型的2倍,用Ag+处理可抑制hps4对缺磷的过敏反应,而用AVG处理则不能,表明hps4中增强的乙烯信号促进了根尖生长素的生物合成,而生长素过量累积抑制了主根的生长[56].在低钾条件下,乙烯促进生长素转运蛋白基因PIN3(PIN-FORMED 3)和PIN7的表达,导致生长素输出量增加,从而阻止驱动侧根形成所需的生长素的局部积累,最终抑制侧根发育[57].此外,在低铁条件下,乙烯和生长素协同抑制根的伸长,并拮抗调控侧根的形成:生长素诱导侧根发育,而乙烯抑制侧根发育[57-58].缺硼(B)会导致乙烯和生长素介导的主根生长受到抑制,其机制可能是缺硼诱导ACS11基因表达,导致乙烯含量增加,而乙烯通过促进生长素的生物合成以及调控生长素的运输机制,激活了局部生长素信号转导,最终导致根细胞长度变短,主根生长受到抑制[59-60]. ...
Ethylene upregulates auxin biosynthesis in Arabidopsis seedlings to enhance inhibition of root cell elongation
1
2007
... 乙烯通过促进生长素(auxin, IAA)的生物合成和运输来调节根的发育.在缺磷条件下,拟南芥乙烯组成型响应突变体hps4的主根生长抑制更为严重.研究发现,hps4中生长素生物合成相关基因的表达量增加,其根尖的生长素合成量是野生型的2倍,用Ag+处理可抑制hps4对缺磷的过敏反应,而用AVG处理则不能,表明hps4中增强的乙烯信号促进了根尖生长素的生物合成,而生长素过量累积抑制了主根的生长[56].在低钾条件下,乙烯促进生长素转运蛋白基因PIN3(PIN-FORMED 3)和PIN7的表达,导致生长素输出量增加,从而阻止驱动侧根形成所需的生长素的局部积累,最终抑制侧根发育[57].此外,在低铁条件下,乙烯和生长素协同抑制根的伸长,并拮抗调控侧根的形成:生长素诱导侧根发育,而乙烯抑制侧根发育[57-58].缺硼(B)会导致乙烯和生长素介导的主根生长受到抑制,其机制可能是缺硼诱导ACS11基因表达,导致乙烯含量增加,而乙烯通过促进生长素的生物合成以及调控生长素的运输机制,激活了局部生长素信号转导,最终导致根细胞长度变短,主根生长受到抑制[59-60]. ...
The Arabidopsis ethylene/jasmonic acid-NRT signaling module coordinates nitrate reallocation and the trade-off between growth and environmental adaptation
2
2014
... 营养胁迫能够分别激活乙烯和茉莉酸(jasmonic acid, JA)信号通路,2个信号汇集到EIN3/EIL1上.EIN3/EIL1一方面调节ORA59(OCTADECANOID-RESPONSIVE ARABIDOPSIS AP2/ERF59)的表达,通过与NRT1.8的启动子结合上调其表达,另一方面通过与未知蛋白一起结合NRT1.5启动子下调其表达[61].除此之外,JA信号通路也通过蛋白COI1(coronatine-insensitive 1)参与NRT1.5的下调.因此,乙烯和JA信号通路通过协同上调NRT1.8和下调NRT1.5来调节胁迫引起的根系硝酸盐的再分配,从而实现胁迫耐受性和植物生长之间的平衡[61]. ...
... [61]. ...
The role of protein turnover in ethylene biosynthesis and response
1
2008
... 乙烯的信号转导及功能受到泛素-蛋白酶体系统的调节[62].ACS作为乙烯生物合成的关键限速酶,其蛋白丰度受到ETO1(ethylene overproducing 1)及其同源蛋白EOL1(ETO1-like)和EOL2的调控[63].ETO1/EOLs是CULLIN3 E3泛素连接酶的组分,可以特异性识别2型ACS并通过26S蛋白酶体进行快速降解[63].14-3-3蛋白是真核生物中高度保守的二聚体蛋白,通过磷酸化介导与靶蛋白相互作用参与多种生物过程[64].在碳饥饿条件下,拟南芥出现叶片黄化衰老的表型,同时,碳饥饿会激活油菜素甾醇(brassinosteroid, BR)信号通路,由BR诱导的磷酸化的SINAT2(seven-in-absentia of A. thaliana 2)和ETO1/EOL2将14-3-3蛋白招募到复合物中,激活SINAT2与EOL2的降解,增加ACS5蛋白的丰度和自噬活性,最终促进乙烯生物合成以响应碳饥饿[65].24-表油菜素内酯(24-epibrassinolide, EBR)是BR家族中的一种.有报道发现,EBR可增加富铁幼苗根系高铁还原酶FRO的活性,却降低了缺铁幼苗中FRO的活性;进一步研究发现,EBR抑制缺铁诱导的乙烯产生,但促进富铁幼苗中乙烯的产生,说明BR可能通过与乙烯交互负调控FRO活性,从而参与植物对缺铁的响应[66]. ...
New insights into the protein turnover regulation in ethylene biosynthesis
2
2015
... 乙烯的信号转导及功能受到泛素-蛋白酶体系统的调节[62].ACS作为乙烯生物合成的关键限速酶,其蛋白丰度受到ETO1(ethylene overproducing 1)及其同源蛋白EOL1(ETO1-like)和EOL2的调控[63].ETO1/EOLs是CULLIN3 E3泛素连接酶的组分,可以特异性识别2型ACS并通过26S蛋白酶体进行快速降解[63].14-3-3蛋白是真核生物中高度保守的二聚体蛋白,通过磷酸化介导与靶蛋白相互作用参与多种生物过程[64].在碳饥饿条件下,拟南芥出现叶片黄化衰老的表型,同时,碳饥饿会激活油菜素甾醇(brassinosteroid, BR)信号通路,由BR诱导的磷酸化的SINAT2(seven-in-absentia of A. thaliana 2)和ETO1/EOL2将14-3-3蛋白招募到复合物中,激活SINAT2与EOL2的降解,增加ACS5蛋白的丰度和自噬活性,最终促进乙烯生物合成以响应碳饥饿[65].24-表油菜素内酯(24-epibrassinolide, EBR)是BR家族中的一种.有报道发现,EBR可增加富铁幼苗根系高铁还原酶FRO的活性,却降低了缺铁幼苗中FRO的活性;进一步研究发现,EBR抑制缺铁诱导的乙烯产生,但促进富铁幼苗中乙烯的产生,说明BR可能通过与乙烯交互负调控FRO活性,从而参与植物对缺铁的响应[66]. ...
... [63].14-3-3蛋白是真核生物中高度保守的二聚体蛋白,通过磷酸化介导与靶蛋白相互作用参与多种生物过程[64].在碳饥饿条件下,拟南芥出现叶片黄化衰老的表型,同时,碳饥饿会激活油菜素甾醇(brassinosteroid, BR)信号通路,由BR诱导的磷酸化的SINAT2(seven-in-absentia of A. thaliana 2)和ETO1/EOL2将14-3-3蛋白招募到复合物中,激活SINAT2与EOL2的降解,增加ACS5蛋白的丰度和自噬活性,最终促进乙烯生物合成以响应碳饥饿[65].24-表油菜素内酯(24-epibrassinolide, EBR)是BR家族中的一种.有报道发现,EBR可增加富铁幼苗根系高铁还原酶FRO的活性,却降低了缺铁幼苗中FRO的活性;进一步研究发现,EBR抑制缺铁诱导的乙烯产生,但促进富铁幼苗中乙烯的产生,说明BR可能通过与乙烯交互负调控FRO活性,从而参与植物对缺铁的响应[66]. ...




{kind=link}
{kind=link}
{kind=link}
{kind=link}