| Special Topic: Insect Physiology and Biochemistry & Pest Biological Control |
|


|
|
|
| Changes in phytohormone levels and gene expression of potato plants in response to potato virus Y and a nonvector insect |
Yang GE1( ),Jian ZHONG1,Yadong ZHANG1,Xiaoli HE1,Lining LIU2,Asim MUNAWAR1,Lijuan MAO3,Zengrong ZHU1,Yi XU4,5,Wenwu ZHOU1() ),Jian ZHONG1,Yadong ZHANG1,Xiaoli HE1,Lining LIU2,Asim MUNAWAR1,Lijuan MAO3,Zengrong ZHU1,Yi XU4,5,Wenwu ZHOU1() |
1.Institute of Insect Sciences, College of Agriculture and Biotechnology, Zhejiang University/Zhejiang Provincial Key Laboratory of Biology of Crop Pathogens and Insects/Ministry of Agriculture and Rural Affairs Key Laboratory of Molecular Biology of Crop Pathogens and Insects, Hangzhou 310058, Zhejiang, China
2.College of Plant Protection, China Agricultural University, Beijing 100193, China
3.Analysis Center of Agrobiology and Environmental Sciences (ACAES), Zhejiang University, Hangzhou 310058, Zhejiang, China
4.Department of Plant Pathology, Nanjing Agricultural University, Nanjing 210095, Jiangsu, China
5.Key Laboratory of Plant Immunity, Nanjing Agricultural University, Nanjing 210095, Jiangsu, China |
|
|
|
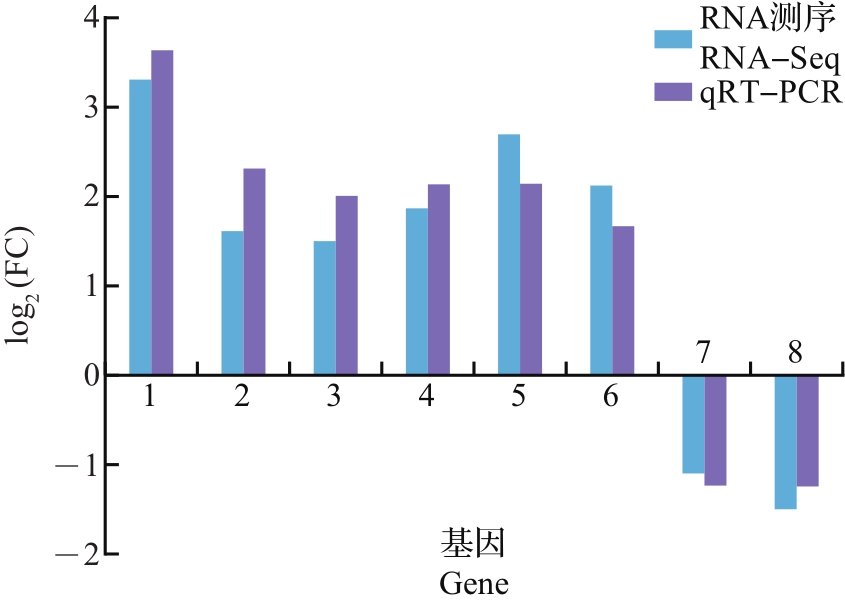
Abstract Potato virusY (PVY) and its nonvector insect of potato tuber moth (Phthorimaea operculella) are important disease and pest in global potato industry. In order to investigate whether PVY infection changed the physiological responses of potato plant to P. operculella, we studied performances of P. operculella on PVY-infected and non-infected healthy plants, and carried out the phytohormone analysis and comparative transcriptomic analysis for potato leaves which were affected by PVY infection and herbiory stimuli (dealing with the oral secretions of potato tuber moth), or co-stressed treatment. The results showed that the resistance to P. operculella was significantly increased in potato plants infected by PVY. Moreover, PVY infection also inhibited the increase of abscisic acid induced by herbivory stimuli. Compared with the healthy control plants, herbivory stimuli and PVY infection treatments produced 3 998 and 104 differentially expressed genes (DEGs), respectively; while 9 178 DEGs were induced by co-stressed treatment, suggesting that the co-stressed treatment may greatly affected the physiological responses in potato plants. Compared with the herbivory stimuli, the co-stressed plants had 743 DEGs, and many of them belong to the transferase activity, the protein processing in endoplasmic reticulum, and the phenylpropanoid biosynthesis pathways. And the expression levels of many genes in abscisic acid synthesis pathway were also down-regulated, which was consisted with the change of this phytohormone. In summary, this study systematically analyzed the physiological responses of potato plant confronted by the single- and co-stressed treatments of PVY infection and herbivory stimuli, which may provide scientific basis for exploring plant-virus-nonvector insect interactions.
|
|
Received: 30 March 2022
Published: 25 April 2023
|
|
|
|
Corresponding Authors:
Wenwu ZHOU
E-mail: 21916167@zju.edu.cn;wenwuzhou@zju.edu.cn
|
|
Cite this article:
Yang GE,Jian ZHONG,Yadong ZHANG,Xiaoli HE,Lining LIU,Asim MUNAWAR,Lijuan MAO,Zengrong ZHU,Yi XU,Wenwu ZHOU. Changes in phytohormone levels and gene expression of potato plants in response to potato virus Y and a nonvector insect. Journal of Zhejiang University (Agriculture and Life Sciences), 2023, 49(2): 167-178.
URL:
https://www.zjujournals.com/agr/10.3785/j.issn.1008-9209.2022.03.301 OR https://www.zjujournals.com/agr/Y2023/V49/I2/167
|
马铃薯应对马铃薯Y病毒与非介体昆虫胁迫的植物激素水平和基因表达变化
马铃薯Y病毒(potato virus Y, PVY)及其非介体昆虫马铃薯块茎蛾(Phthorimaea operculella)是世界马铃薯产业的重要病虫害,为探究PVY是否影响马铃薯对马铃薯块茎蛾的生理响应,本研究分析了马铃薯块茎蛾在PVY侵染和健康对照植株上的生长表现,并对无病虫害、PVY侵染、模拟虫害处理(用马铃薯块茎蛾口腔分泌物涂抹),以及PVY侵染和模拟虫害协同处理的植株叶片进行了植物激素测定和比较转录组分析。结果显示:马铃薯感染PVY后对马铃薯块茎蛾的抗性显著提高;PVY侵染可抑制模拟虫害造成的脱落酸含量上升;与健康对照组相比,模拟虫害和PVY侵染处理分别诱导产生3 998个和104个差异表达基因,而两者协同处理可诱导产生9 178个差异表达基因,说明这2种胁迫共存对马铃薯产生了极大的生理影响。与模拟虫害处理相比,PVY侵染和模拟虫害协同处理诱导产生743个差异表达基因,而这些基因主要富集在转移酶活性、内质网中蛋白质加工、苯丙素生物合成等通路中,而脱落酸合成通路中众多基因表达下调,与该植物激素变化情况一致。综上所述,本研究系统分析了马铃薯应对PVY侵染、马铃薯块茎蛾模拟虫害以及两者协同处理的生理反应,为探究“植物-病毒-非介体昆虫”三者互作提供了科学依据。
关键词:
马铃薯,
马铃薯Y病毒,
马铃薯块茎蛾,
协同处理,
生理响应
|
|
| [1] |
NOMAN A, AQEEL M, QASIM M, et al. Plant-insect-microbe interaction: a love triangle between enemies in ecosystem[J]. Science of the Total Environment, 2020, 699: 134181. DOI: 10.1016/j.scitotenv.2019.134181
doi: 10.1016/j.scitotenv.2019.134181
|
|
|
| [2] |
FELTON G W, KORTH K L. Trade-offs between pathogen and herbivore resistance[J]. Current Opinion in Plant Biology, 2000, 3(4): 309-314. DOI: 10.1016/S1369-5266(00)00086-8
doi: 10.1016/S1369-5266(00)00086-8
|
|
|
| [3] |
HOWE G A, JANDER G. Plant immunity to insect herbivores[J]. Annual Review of Plant Biology, 2008, 59: 41-66. DOI: 10.1146/annurev.arplant.59.032607.092825
doi: 10.1146/annurev.arplant.59.032607.092825
|
|
|
| [4] |
AHUJA I, DE VOS R C H, BONES A M, et al. Plant molecular stress responses face climate change[J]. Trends in Plant Science, 2010, 15(12): 664-674. DOI: 10.1016/j.tplants.2010.08.002
doi: 10.1016/j.tplants.2010.08.002
|
|
|
| [5] |
VAN DIJK L J A, EHRLÉN J, TACK A J M. Direct and insect-mediated effects of pathogens on plant growth and fitness[J]. Journal of Ecology, 2021, 109(7): 2769-2779. DOI: 10.1111/1365-2745.13689
doi: 10.1111/1365-2745.13689
|
|
|
| [6] |
DONNELLY R, GILLIGAN C A. What is pathogen-mediated insect superabundance?[J]. Journal of the Royal Society Interface, 2020, 17(170): 20200229. DOI: 10.1098/rsif.2020.0229
doi: 10.1098/rsif.2020.0229
|
|
|
| [7] |
SHIKANO I, ROSA C, TAN C W, et al. Tritrophic interactions: microbe-mediated plant effects on insect herbi-vores[J]. Annual Review of Phytopathology, 2017, 55: 313-331. DOI: 10.1146/annurev-phyto-080516-035319
doi: 10.1146/annurev-phyto-080516-035319
|
|
|
| [8] |
EIGENBRODE S D, DING H J, SHIEL P, et al. Volatiles from potato plants infected with potato leafroll virus attract and arrest the virus vector, Myzus persicae (Homoptera: Aphididae)[J]. Proceedings of the Royal Society B: Biological Sciences, 2002, 269(1490): 455-460. DOI: 10.1098/rspb.2001.1909
doi: 10.1098/rspb.2001.1909
|
|
|
| [9] |
LI R, WELDEGERGIS B T, LI J, et al. Virulence factors of geminivirus interact with MYC2 to subvert plant resistance and promote vector performance[J]. The Plant Cell, 2014, 26(12): 4991-5008. DOI: 10.1105/tpc.114.133181
doi: 10.1105/tpc.114.133181
|
|
|
| [10] |
ABE H, TOMITAKA Y, SHIMODA T, et al. Antagonistic plant defense system regulated by phytohormones assists interactions among vector insect, thrips and a tospovirus[J]. Plant and Cell Physiology, 2012, 53(1): 204-212. DOI: 10.1093/pcp/pcr173
doi: 10.1093/pcp/pcr173
|
|
|
| [11] |
ZHAO P Z, YAO X M, CAI C X, et al. Viruses mobilize plant immunity to deter nonvector insect herbivores[J]. Science Advances, 2019, 5(8): eaav9801. DOI: 10.1126/sciadv.aav9801
doi: 10.1126/sciadv.aav9801
|
|
|
| [12] |
KERSCH-BECKER M F, THALER J S. Virus strains differentially induce plant susceptibility to aphid vectors and chewing herbivores[J]. Oecologia, 2014, 174(3): 883-892. DOI: 10.1007/s00442-013-2812-7
doi: 10.1007/s00442-013-2812-7
|
|
|
| [13] |
PETEK M, ROTTER A, KOGOVŠEK P, et al. Potato virus Y infection hinders potato defence response and renders plants more vulnerable to Colorado potato beetle attack[J]. Molecular Ecology, 2014, 23(21): 5378-5391. DOI: 10.1111/mec.12932
doi: 10.1111/mec.12932
|
|
|
| [14] |
BOSTOCK R M, KARBAN R, THALER J S, et al. Signal interactions in induced resistance to pathogens and insect herbivores[J]. European Journal of Plant Pathology, 2001, 107: 103-111. DOI: 10.1023/A:1008703904253
doi: 10.1023/A:1008703904253
|
|
|
| [15] |
XU J, WANG X J, ZU H Y, et al. Molecular dissection of rice phytohormone signaling involved in resistance to a piercing-sucking herbivore[J]. New Phytologist, 2021, 230(4): 1639-1652. DOI: 10.1111/nph.17251
doi: 10.1111/nph.17251
|
|
|
| [16] |
MAYERS C N, LEE K C, MOORE C A, et al. Salicylic acid-induced resistance to Cucumber mosaic virus in squash and Arabidopsis thaliana: contrasting mechanisms of induction and antiviral action[J]. Molecular Plant-Microbe Interaction, 2005, 18(5): 428-434. DOI: 10.1094/MPMI-18-0428
doi: 10.1094/MPMI-18-0428
|
|
|
| [17] |
SONG S S, HUANG H, GAO H, et al. Interaction between MYC2 and ETHYLENE INSENSITIVE3 modulates antagonism between jasmonate and ethylene signaling in Arabidopsis [J]. The Plant Cell, 2014, 26(1): 263-279. DOI: 10.1105/tpc.113.120394
doi: 10.1105/tpc.113.120394
|
|
|
| [18] |
HILLWIG M S, CHIOZZA M, CASTEEL C L, et al. Abscisic acid deficiency increases defence responses against Myzus persicae in Arabidopsis [J]. Molecular Plant Pathology, 2016, 17(2): 225-235. DOI: 10.1111/mpp.12274
doi: 10.1111/mpp.12274
|
|
|
| [19] |
CUI H Y, SUN Y C, CHEN F J, et al. Elevated O3 and TYLCV infection reduce the suitability of tomato as a host for the whitefly Bemisia tabaci [J]. International Journal of Molecular Sciences, 2016, 17(12): 1964. DOI: 10.3390/ijms17121964
doi: 10.3390/ijms17121964
|
|
|
| [20] |
GLAIS L, TRIBODE M, KERLAN C. Specific detection of the PVYN-Wvariant of Potato virus Y [J]. Journal of Virological Methods, 2005, 125(2): 131-136. DOI: 10.1016/j.jviromet.2005.01.007
doi: 10.1016/j.jviromet.2005.01.007
|
|
|
| [21] |
LI W B, ZARKA K A, DOUCHES D S, et al. Coexpression of potato PVYo coat protein and cryV-Bt genes in potato[J]. Journal of the American Society for Horticultural Science, 1999, 124(3): 218-223. DOI: 10.21273/JASHS.124.3.218
doi: 10.21273/JASHS.124.3.218
|
|
|
| [22] |
SHRESTHA D, WENNINGER E J, HUTCHINSON P J S, et al. Interactions among potato genotypes,growth stages, virus strains, and inoculation methods in the Potato virus Y and green peach aphid pathosystem[J]. Environmental Entomology, 2014, 43(3): 662-671. DOI: 10.1603/EN13323
doi: 10.1603/EN13323
|
|
|
| [23] |
CHEN C Y, LIU Y Q, SONG W M, et al. An effector from cotton bollworm oral secretion impairs host plant defense signaling[J]. PNAS, 2019, 116(28): 14331-14338. DOI: 10.1073/pnas.1905471116
doi: 10.1073/pnas.1905471116
|
|
|
| [24] |
ZHANG C P, LI J, LI S, et al. ZmMPK6 and ethylene signaling negatively regulate the accumulation of anti-insect metabolites DIMBOA and DIMBOA-Glcinmaize inbred line A188[J]. New Phytologist, 2021, 229(4): 2273-2287. DOI: 10.1111/nph.16974
doi: 10.1111/nph.16974
|
|
|
| [25] |
蔡雅洁.马铃薯应答虫害胁迫的蛋白组分析及关键基因的功能研究[D].浙江,杭州:浙江大学,2021:12-15.
CAI Y J. Proteomic analysis od potato defense response to herbivore stress and functional study of key regulatory genes[D]. Hangzhou, Zhejiang: Zhejiang University, 2021: 12-15. (in Chinese with English abstract)
|
|
|
| [26] |
梁承娟.马铃薯受块茎蛾胁迫的转录组分析及马铃薯基因编辑体系的初步构建[D].福建,福州:福建农林大学,2021:10-15.
LIANG C J. Transcriptome analysis of potato leaf damaged by potato tuber moth and preliminary construction of gene editing system in potato[D]. Fuzhou, Fujian: Fujian Agriculture and Forestry University, 2021: 10-15. (in Chinese with English abstract)
|
|
|
| [27] |
LI R, ZHANG J, LI J C, et al. Prioritizing plant defence over growth through WRKY regulation facilitates infestation by non-target herbivores[J]. eLife, 2015, 4: e04805. DOI: 10.7554/eLife.04805
doi: 10.7554/eLife.04805
|
|
|
| [28] |
PAN L L, MIAO H Y, WANG Q M, et al. Virus induced phytohormone dynamics and their effects on plant-insect interactions[J]. New Phytologist, 2021, 230(4): 1305-1320. DOI: 10.1111/nph.17261
doi: 10.1111/nph.17261
|
|
|
| [29] |
PIETERSE C M J, VAN DER DOES D, ZAMIOUDIS C, et al. Hormonal modulation of plant immunity[J]. Annual Review of Cell & Developmental Biology, 2012, 28: 489-521. DOI: 10.1146/annurev-cellbio-092910-154055
doi: 10.1146/annurev-cellbio-092910-154055
|
|
|
| [30] |
王金秋,武舜臣.马铃薯主粮化战略的动力、障碍与前景[J].农业经济,2018(4):17-19. DOI:10.3969/j.issn.1001-6139.2018.04.006
WANG J Q, WU S C. The driving force, obstacle and prospect of potato staple food strategy[J]. Agricultural Economy, 2018(4): 17-19. (in Chinese)
doi: 10.3969/j.issn.1001-6139.2018.04.006
|
|
|
| [31] |
SUN Y C, GUO H J, YUAN L, et al. Plant stomatal closure improves aphid feeding under elevated CO2 [J]. Global Change Biology, 2015, 21(7): 2739-2748. DOI: 10.1111/gcb.12858
doi: 10.1111/gcb.12858
|
|
|
| [32] |
BAK A, PATTON M F, PERILLA-HENAO L M, et al. Ethylene signaling mediates potyvirus spread by aphid vectors[J]. Oecologia, 2019, 190: 139-148. DOI: 10.1007/s00442-019-04405-0
doi: 10.1007/s00442-019-04405-0
|
|
|
| [33] |
WU J Q, HETTENHAUSEN C, MELDAU S, et al. Herbivory rapidly activates MAPK signaling in attacked and unattacked leaf regions but not between leaves of Nicotiana attenuata [J]. The Plant Cell, 2007, 19(3): 1096-1122. DOI: 10.1105/tpc.106.049353
doi: 10.1105/tpc.106.049353
|
|
|
| [34] |
KANDOTH P K, RANF S, PANCHOLI S S, et al. Tomato MAPKs LeMPK1, LeMPK2, and LeMPK3 function in the systemin-mediated defense response against herbivorous insects[J]. PNAS, 2007, 104(29): 12205-12210. DOI: 10.1073/pnas.0700344104
doi: 10.1073/pnas.0700344104
|
|
|
|
Viewed |
|
|
|
Full text
|
|
|
|
|
Abstract
|
|
|
|
|
Cited |
|
|
|
|
| |
Shared |
|
|
|
|
| |
Discussed |
|
|
|
|





{kind=link}
{kind=link}