| Animal sciences & veterinary medicines |
|

|
|
|
| Soluble expression of nucleocapsid protein of feline coronavirus and reactivity identification of its polyclonal antibody |
Dan ZHANG1( ),Shuting LU1(),Chenhe LU1,Ziyi WANG1,Lihua XU1,2,Yixuan CHEN1,Shengwen WANG1,Zi’an JIN1,3,Chengzhang NI1,Jiyong ZHOU1,Xiaojuan ZHENG1() ),Shuting LU1(),Chenhe LU1,Ziyi WANG1,Lihua XU1,2,Yixuan CHEN1,Shengwen WANG1,Zi’an JIN1,3,Chengzhang NI1,Jiyong ZHOU1,Xiaojuan ZHENG1() |
1.Key Laboratory of Animal Virology, Ministry of Agriculture and Rural Affairs, Center for Veterinary Sciences, Zhejiang University, Hangzhou 310058, Zhejiang, China
2.Institute of Animal Husbandry and Veterinary Science, Zhejiang Academy of Agricultural Sciences, Hangzhou 310021, Zhejiang, China
3.Hainan Institute of Zhejiang University, Sanya 572025, Hainan, China |
|
|
|
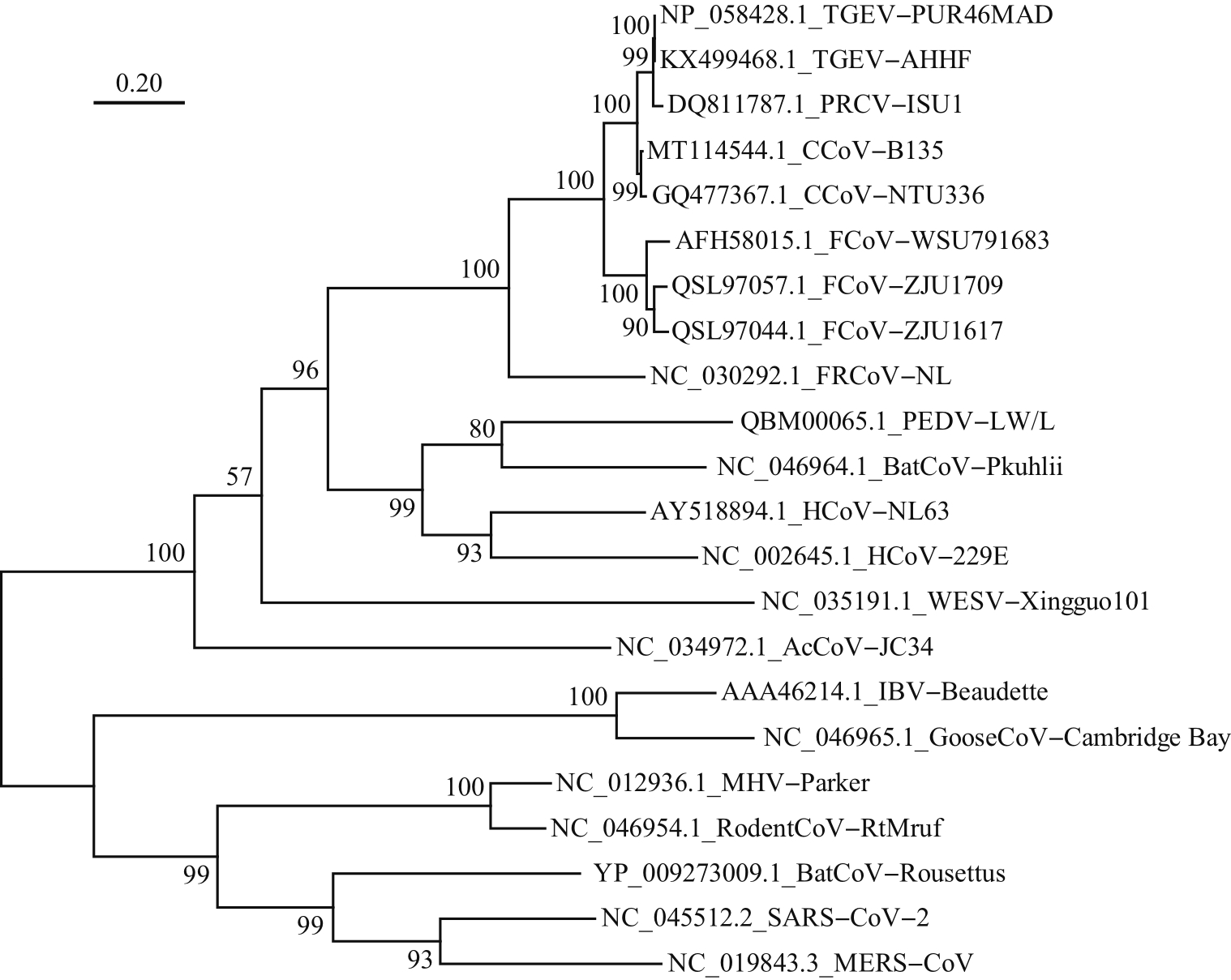
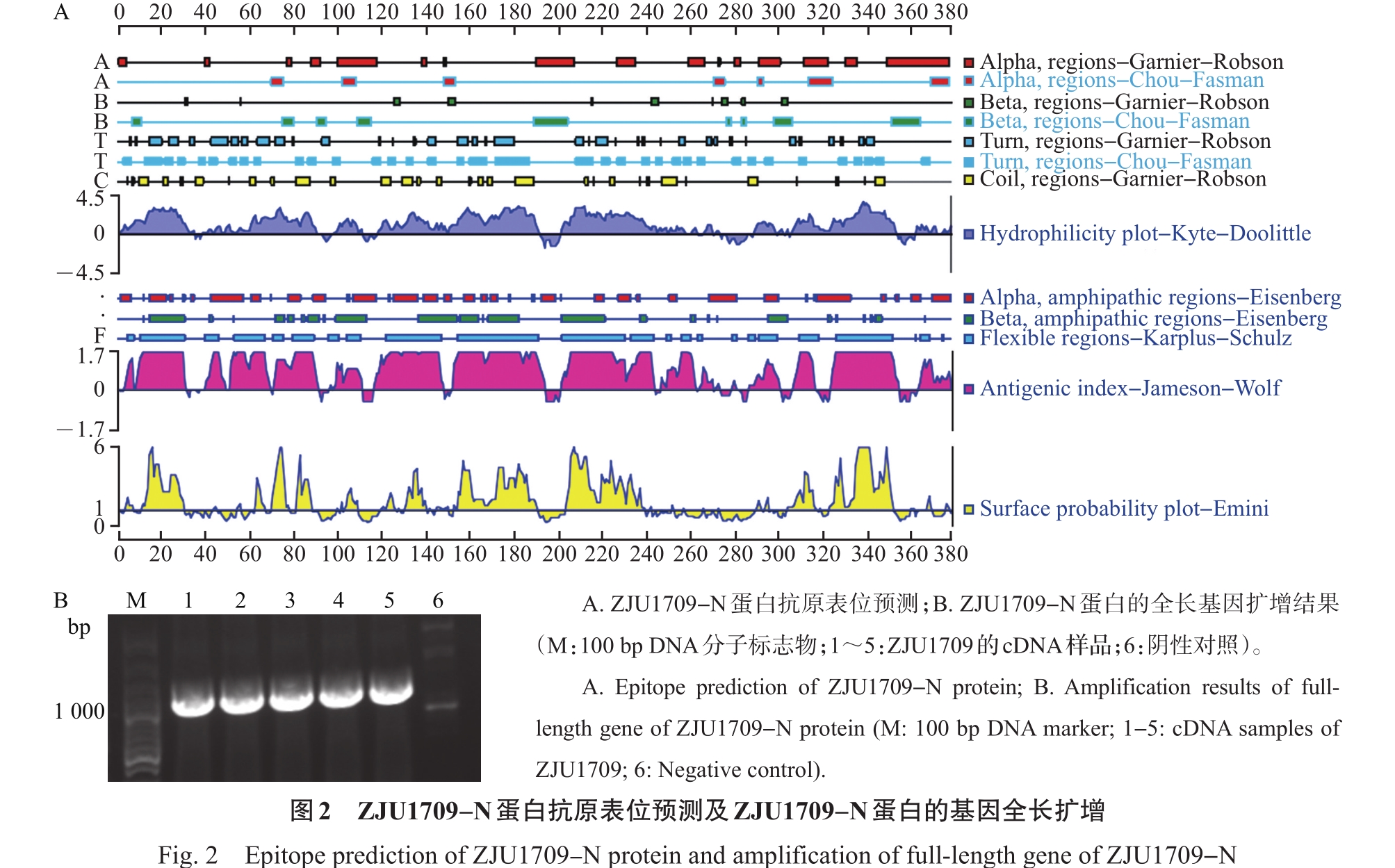
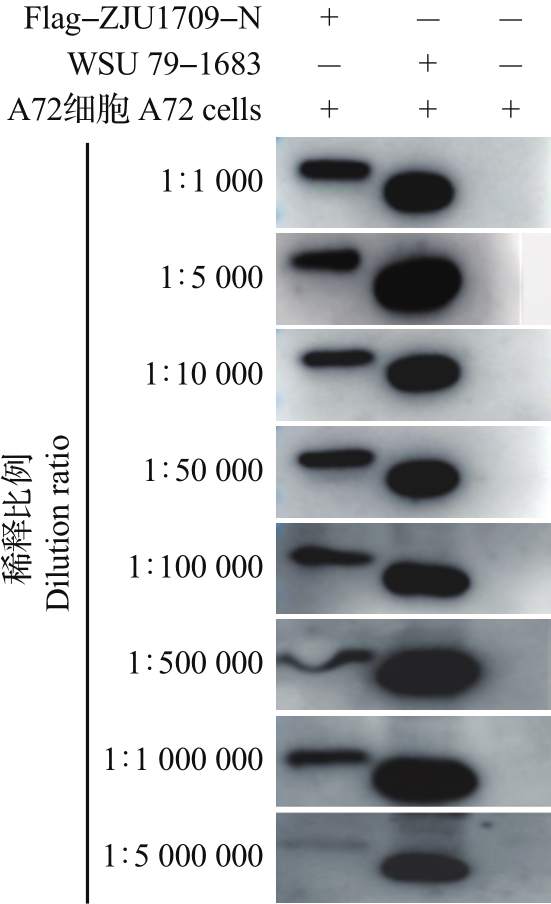
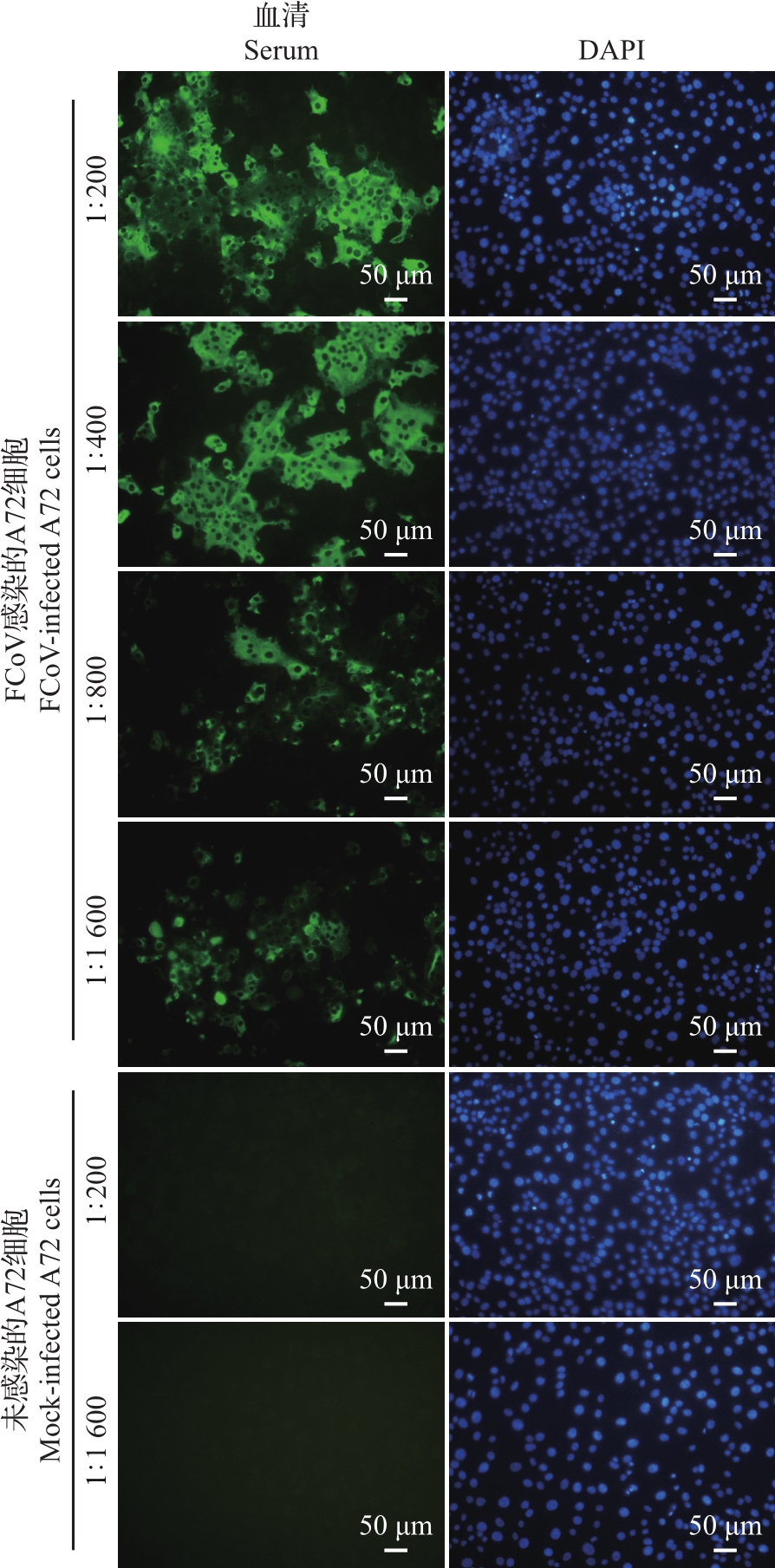
Abstract To prepare the specific antibodies against nucleocapsid (N) protein of feline coronavirus (FCoV), the full-length cDNA of N gene was amplified from type ⅠFCoV (ZJU1709) and subcloned into pCold TF prokaryotic expression vector. The recombinant N protein was induced by isopropyl-β-D-thiogalactopyranoside (IPTG) at low temperature and affinity-purified using nickel column. The rabbit polyclonal antiserum was prepared using the purified recombinant N protein as antigen. Finally, the enzyme-linked immunosorbent assay (ELISA), Western blotting (WB) and indirect immunofluorescence assay (IFA) were applied to identify the reactivity of rabbit polyclonal antiserum. The results showed that the soluble recombinant N protein with a molecular weight of about 100 kDa was successfully induced and further purified under non-denaturing condition at a concentration of 1.4 mg/mL. The ELISA titer of the prepared rabbit polyclonal antiserum of N protein can reach 1×106, and it can react with the recombinant Flag-N eukaryotic protein and N protein in FCoV-infected cells by WB and IFA. Cross-reactivity analysis showed that the polyclonal antibody can react with N proteins of alphacoronavirus, including various subtypes of FCoV, as well as canine coronavirus (CCoV), porcine transmissible gastroenteritis virus (TGEV), and porcine epidemic diarrhea virus (PEDV), but failed to react with N proteins of betacoronavirus of severe acute respiratory syndrome-related coronavirus 2 (SARS-CoV-2) and gammacoronavirus of infectious bronchitis virus (IBV). In summary, the soluble recombinant N protein expressed by using pCold TF prokaryotic system showed strong immunogenicity. The rabbit polyclonal antiserum against N protein of FCoV can recognize N proteins of multiple species from alphacoronavirus, but showed no cross-reactivity with N proteins of betacoronavirus and gammacoronavirus. This antibody facilitates to study the replication mechanism of FCoV and develop efficient methods for antigen or antibody detection.
|
|
Received: 07 May 2021
Published: 25 June 2023
|
|
|
|
Corresponding Authors:
Xiaojuan ZHENG
E-mail: zhangdan9395@163.com;21717092@zju.edu.cn;zhengxiaojuan@zju.edu.cn
|
|
Cite this article:
Dan ZHANG,Shuting LU,Chenhe LU,Ziyi WANG,Lihua XU,Yixuan CHEN,Shengwen WANG,Zi’an JIN,Chengzhang NI,Jiyong ZHOU,Xiaojuan ZHENG. Soluble expression of nucleocapsid protein of feline coronavirus and reactivity identification of its polyclonal antibody. Journal of Zhejiang University (Agriculture and Life Sciences), 2023, 49(3): 424-434.
URL:
https://www.zjujournals.com/agr/10.3785/j.issn.1008-9209.2021.05.072 OR https://www.zjujournals.com/agr/Y2023/V49/I3/424
|
猫冠状病毒核衣壳蛋白的可溶性表达与多克隆抗体反应性鉴定
为制备猫冠状病毒(feline coronavirus, FCoV)核衣壳(nucleocapsid, N)蛋白的特异性抗体,本研究将Ⅰ型FCoV(ZJU1709)的N蛋白基因全长cDNA亚克隆到pCold TF原核表达载体上,通过低温诱导表达、镍柱亲和纯化获得可溶性重组N蛋白;进一步以纯化的重组N蛋白为免疫原制备兔多克隆抗血清(多抗血清),再通过酶联免疫吸附测定(enzyme-linked immunosorbent assay, ELISA)、蛋白质印迹法(Western blotting, WB)、间接免疫荧光试验(indirect immunofluorescence assay, IFA)等多种方法对兔多抗血清进行反应性鉴定。结果显示:经异丙基-β-D-硫代半乳糖苷(isopropyl-β-D-thiogalactopyranoside, IPTG)和低温诱导,成功表达出分子量约为100 kDa的可溶性重组N蛋白,在非变性条件下亲和纯化获得的重组N蛋白质量浓度达到1.4 mg/mL;N蛋白兔多抗血清的ELISA效价可达1×106,与真核表达的重组蛋白Flag-N以及FCoV感染细胞中的N蛋白均具有良好的WB和IFA反应性。交叉反应性结果显示:该多克隆抗体能与α属冠状病毒中不同亚型的FCoV以及犬冠状病毒(canine coronavirus, CCoV)、猪传染性胃肠炎病毒(porcine transmissible gastroenteritis virus, TGEV)、猪流行性腹泻病毒(porcine epidemic diarrhea virus, PEDV)的N蛋白发生反应,但不与β属冠状病毒中的严重急性呼吸综合征冠状病毒2(severe acute respiratory syndrome-related coronavirus 2, SARS-CoV-2)和γ属冠状病毒中的传染性支气管炎病毒(infectious bronchitis virus, IBV)的N蛋白发生反应。综上所述,以pCold TF原核系统表达的可溶性重组N蛋白具有良好的免疫原性,以该蛋白制备的FCoV N蛋白兔多抗血清能识别多种来自不同物种的α属冠状病毒N蛋白,但与β、γ属冠状病毒N蛋白没有交叉反应性,这为进一步研究FCoV复制机制以及开发高效的抗原和抗体检测技术奠定了基础。
关键词:
猫冠状病毒,
核衣壳蛋白,
可溶性表达,
多克隆抗体,
交叉反应性
|
|
| [1] |
SHI J Z, WEN Z Y, ZHONG G X, et al. Susceptibility of ferrets, cats, dogs, and other domesticated animals to SARS-coronavirus 2[J]. Science, 2020, 368(6494): 1016-1021. DOI: 10.1126/science.abb7015
doi: 10.1126/science.abb7015
|
|
|
| [2] |
JACKWOOD M W. Review of infectious bronchitis virus around the world[J]. Avian Diseases, 2012, 56(4): 634-641. DOI: 10.1637/10227-043012-Review.1
doi: 10.1637/10227-043012-Review.1
|
|
|
| [3] |
LEDNICKY J A, TAGLIAMONTE M S, WHITE S K, et al. Independent infections of porcine deltacoronavirus among haitian children[J]. Nature, 2021, 600(7887): 133-137. DOI: 10.1038/s41586-021-04111-z
doi: 10.1038/s41586-021-04111-z
|
|
|
| [4] |
LI W T, HULSWIT R J G, KENNEY S P, et al. Broad receptor engagement of an emerging global coronavirus may potentiate its diverse cross-species transmissibility[J]. PNAS, 2018, 115(22): E5135-E5143. DOI: 10.1073/pnas.1802879115
doi: 10.1073/pnas.1802879115
|
|
|
| [5] |
TRESNAN D B, LEVIS R, HOLMES K V. Feline amino-peptidase N serves as a receptor for feline, canine, porcine, and human coronaviruses in serogroup Ⅰ[J]. Journal of Virology, 1996, 70(12): 8669-8674. DOI: 10.1128/jvi.70.12.8669-8674.1996
doi: 10.1128/jvi.70.12.8669-8674.1996
|
|
|
| [6] |
TUSELL S M, SCHITTONE S A, HOLMES K V. Mutational analysis of aminopeptidase N, a receptor for several group 1 coronaviruses, identifies key determinants of viral host range[J]. Journal of Virology, 2007, 81(3): 1261-1273. DOI: 10.1128/jvi.01510-06
doi: 10.1128/jvi.01510-06
|
|
|
| [7] |
HERREWEGH A A P M, SMEENK I, HORZINEK M C, et al. Feline coronavirus type Ⅱ strains 79-1683 and 79-1146 originate from a double recombination between feline corona-virus type Ⅰ and canine coronavirus[J]. Journal of Virology, 1998, 72(5): 4508-4514. DOI: 10.1128/jvi.72.5.4508-4514.1998
doi: 10.1128/jvi.72.5.4508-4514.1998
|
|
|
| [8] |
STOUT A E, ANDRÉ N M, WHITTAKER G R. Feline coronavirus and feline infectious peritonitis in nondomestic felid species[J]. Journal of Zoo and Wildlife Medicine, 2021, 52(1): 14-27. DOI: 10.1638/2020-0134
doi: 10.1638/2020-0134
|
|
|
| [9] |
YANG T J, CHANG Y C, KO T P, et al. Cryo-EM analysis of a feline coronavirus spike protein reveals a unique structure and camouflaging glycans[J]. PNAS, 2020, 117(3): 1438-1446. DOI: 10.1073/pnas.1908898117
doi: 10.1073/pnas.1908898117
|
|
|
| [10] |
MCBRIDE R, VAN ZYL M, FIELDING B C. The coronavirus nucleocapsid is a multifunctional protein[J]. Viruses, 2014, 6(8): 2991-3018. DOI: 10.3390/v6082991
doi: 10.3390/v6082991
|
|
|
| [11] |
HURST K R, KOETZNER C A, MASTERS P S. Charac-terization of a critical interaction between the coronavirus nucleocapsid protein and nonstructural protein 3 of the viral replicase-transcriptase complex[J]. Journal of Virology, 2013, 87(16): 9159-9172. DOI: 10.1128/jvi.01275-13
doi: 10.1128/jvi.01275-13
|
|
|
| [12] |
TAKANO T, ISHIHARA Y, MATSUOKA M, et al. Use of recombinant nucleocapsid proteins for serological diagnosis of feline coronavirus infection by three immunochromato-graphic tests[J]. Journal of Virology Methods, 2014, 196: 1-6. DOI: 10.1016/j.jviromet.2013.10.014
doi: 10.1016/j.jviromet.2013.10.014
|
|
|
|
Viewed |
|
|
|
Full text
|
|
|
|
|
Abstract
|
|
|
|
|
Cited |
|
|
|
|
| |
Shared |
|
|
|
|
| |
Discussed |
|
|
|
|