Coastal blue carbon in China as a nature-based solution toward carbon neutrality
1
2023
... 为了实现《巴黎协定》设定的“到本世纪末全球平均气温的上升幅度控制在1.5 ℃以内”的目标,必须在2050年达到二氧化碳(CO2)净零排放.固定和储存的碳被称为“蓝碳”.在全球气候变化的背景下,滨海蓝碳生态系统因其巨大的碳汇潜力而受到广泛关注.滨海蓝碳生态系统主要包括红树林、盐沼、海草床三大传统蓝碳生态系统和海藻场这一新兴蓝碳生态系统,如图1所示.其中三大传统蓝碳生态系统贡献了全球海洋沉积物中超过60%的有机碳,而总面积只占海洋生态系统的0.5%[1].滨海蓝碳生态系统每年的CO2封存量达到2.2×104 Mg/hm2,是陆地森林生态系统的数十倍乃至数百倍[2].因此,滨海蓝碳生态系统作为重要的天然碳汇,在全球碳循环和缓解气候变化中发挥着至关重要的作用. ...
A blueprint for blue carbon: Toward an improved understanding of the role of vegetated coastal habitats in sequestering CO2
1
2011
... 为了实现《巴黎协定》设定的“到本世纪末全球平均气温的上升幅度控制在1.5 ℃以内”的目标,必须在2050年达到二氧化碳(CO2)净零排放.固定和储存的碳被称为“蓝碳”.在全球气候变化的背景下,滨海蓝碳生态系统因其巨大的碳汇潜力而受到广泛关注.滨海蓝碳生态系统主要包括红树林、盐沼、海草床三大传统蓝碳生态系统和海藻场这一新兴蓝碳生态系统,如图1所示.其中三大传统蓝碳生态系统贡献了全球海洋沉积物中超过60%的有机碳,而总面积只占海洋生态系统的0.5%[1].滨海蓝碳生态系统每年的CO2封存量达到2.2×104 Mg/hm2,是陆地森林生态系统的数十倍乃至数百倍[2].因此,滨海蓝碳生态系统作为重要的天然碳汇,在全球碳循环和缓解气候变化中发挥着至关重要的作用. ...
Future carbon emissions from global mangrove forest loss
1
2021
... 滨海蓝碳增汇技术是指增加易于管理的滨海蓝碳生态系统中由海洋生物驱动的碳通量及碳储量的相关技术.目前滨海蓝碳增汇的主要途径是红树林、盐沼、海草床、海藻场生态系统的保护与修复.全球红树林每年的CO2封存量为(1.7±0.2)Mg/hm2,红树林面积约为1.5×107 hm2,占世界森林总面积的0.4%;然而,在过去30年间,全球红树林面积约减少了1.0×106 hm2,预计到2100年末,因红树林减少导致的CO2排放量为2.4×109~3.4×109 Mg[3].全球盐沼的碳储量为(334.4±3.5)Mg/hm2,盐沼面积约为5.5×106 hm2,占滨海蓝碳生态系统总面积的4%[4];2000—2019年,全球盐沼面积总损失量约为1.4×105 hm2,相当于新加坡面积的2倍,这导致每年约增加了1.6×107 Mg的CO2排放量[5].全球海草床储存的有机碳约达2.0×1010 Mg,海草床面积约为7.9×107 hm2,占全球海洋面积的2%;自20世纪以来,全球海草床的年平均损失率为1.5%,导致每年有1.1×107~2.3×107 Mg有机碳加速分解[6].大型海藻是海岸带的主要初级生产者,其净初级生产力为1.0×109~2.0×109 Mg/a,海藻场面积为2.8×108~4.3×108 hm2[7];在过去50年间,由于海洋污染、捕捞、物种入侵、水产养殖以及全球变暖等原因,约60%的海藻场已退化[8].滨海蓝碳生态系统的退化显著削弱了其碳汇能力,因此,亟须修复滨海蓝碳生态系统,提高其碳汇能力,这对推动“碳达峰”和“碳中和”进程具有重要意义. ...
Quantification of blue carbon in tropical salt marshes and their role in climate change mitigation
1
2022
... 滨海蓝碳增汇技术是指增加易于管理的滨海蓝碳生态系统中由海洋生物驱动的碳通量及碳储量的相关技术.目前滨海蓝碳增汇的主要途径是红树林、盐沼、海草床、海藻场生态系统的保护与修复.全球红树林每年的CO2封存量为(1.7±0.2)Mg/hm2,红树林面积约为1.5×107 hm2,占世界森林总面积的0.4%;然而,在过去30年间,全球红树林面积约减少了1.0×106 hm2,预计到2100年末,因红树林减少导致的CO2排放量为2.4×109~3.4×109 Mg[3].全球盐沼的碳储量为(334.4±3.5)Mg/hm2,盐沼面积约为5.5×106 hm2,占滨海蓝碳生态系统总面积的4%[4];2000—2019年,全球盐沼面积总损失量约为1.4×105 hm2,相当于新加坡面积的2倍,这导致每年约增加了1.6×107 Mg的CO2排放量[5].全球海草床储存的有机碳约达2.0×1010 Mg,海草床面积约为7.9×107 hm2,占全球海洋面积的2%;自20世纪以来,全球海草床的年平均损失率为1.5%,导致每年有1.1×107~2.3×107 Mg有机碳加速分解[6].大型海藻是海岸带的主要初级生产者,其净初级生产力为1.0×109~2.0×109 Mg/a,海藻场面积为2.8×108~4.3×108 hm2[7];在过去50年间,由于海洋污染、捕捞、物种入侵、水产养殖以及全球变暖等原因,约60%的海藻场已退化[8].滨海蓝碳生态系统的退化显著削弱了其碳汇能力,因此,亟须修复滨海蓝碳生态系统,提高其碳汇能力,这对推动“碳达峰”和“碳中和”进程具有重要意义. ...
Global hotspots of salt marsh change and carbon emissions
1
2022
... 滨海蓝碳增汇技术是指增加易于管理的滨海蓝碳生态系统中由海洋生物驱动的碳通量及碳储量的相关技术.目前滨海蓝碳增汇的主要途径是红树林、盐沼、海草床、海藻场生态系统的保护与修复.全球红树林每年的CO2封存量为(1.7±0.2)Mg/hm2,红树林面积约为1.5×107 hm2,占世界森林总面积的0.4%;然而,在过去30年间,全球红树林面积约减少了1.0×106 hm2,预计到2100年末,因红树林减少导致的CO2排放量为2.4×109~3.4×109 Mg[3].全球盐沼的碳储量为(334.4±3.5)Mg/hm2,盐沼面积约为5.5×106 hm2,占滨海蓝碳生态系统总面积的4%[4];2000—2019年,全球盐沼面积总损失量约为1.4×105 hm2,相当于新加坡面积的2倍,这导致每年约增加了1.6×107 Mg的CO2排放量[5].全球海草床储存的有机碳约达2.0×1010 Mg,海草床面积约为7.9×107 hm2,占全球海洋面积的2%;自20世纪以来,全球海草床的年平均损失率为1.5%,导致每年有1.1×107~2.3×107 Mg有机碳加速分解[6].大型海藻是海岸带的主要初级生产者,其净初级生产力为1.0×109~2.0×109 Mg/a,海藻场面积为2.8×108~4.3×108 hm2[7];在过去50年间,由于海洋污染、捕捞、物种入侵、水产养殖以及全球变暖等原因,约60%的海藻场已退化[8].滨海蓝碳生态系统的退化显著削弱了其碳汇能力,因此,亟须修复滨海蓝碳生态系统,提高其碳汇能力,这对推动“碳达峰”和“碳中和”进程具有重要意义. ...
Estimating global “blue carbon” emissions from conversion and degradation of vegetated coastal ecosystems
1
2012
... 滨海蓝碳增汇技术是指增加易于管理的滨海蓝碳生态系统中由海洋生物驱动的碳通量及碳储量的相关技术.目前滨海蓝碳增汇的主要途径是红树林、盐沼、海草床、海藻场生态系统的保护与修复.全球红树林每年的CO2封存量为(1.7±0.2)Mg/hm2,红树林面积约为1.5×107 hm2,占世界森林总面积的0.4%;然而,在过去30年间,全球红树林面积约减少了1.0×106 hm2,预计到2100年末,因红树林减少导致的CO2排放量为2.4×109~3.4×109 Mg[3].全球盐沼的碳储量为(334.4±3.5)Mg/hm2,盐沼面积约为5.5×106 hm2,占滨海蓝碳生态系统总面积的4%[4];2000—2019年,全球盐沼面积总损失量约为1.4×105 hm2,相当于新加坡面积的2倍,这导致每年约增加了1.6×107 Mg的CO2排放量[5].全球海草床储存的有机碳约达2.0×1010 Mg,海草床面积约为7.9×107 hm2,占全球海洋面积的2%;自20世纪以来,全球海草床的年平均损失率为1.5%,导致每年有1.1×107~2.3×107 Mg有机碳加速分解[6].大型海藻是海岸带的主要初级生产者,其净初级生产力为1.0×109~2.0×109 Mg/a,海藻场面积为2.8×108~4.3×108 hm2[7];在过去50年间,由于海洋污染、捕捞、物种入侵、水产养殖以及全球变暖等原因,约60%的海藻场已退化[8].滨海蓝碳生态系统的退化显著削弱了其碳汇能力,因此,亟须修复滨海蓝碳生态系统,提高其碳汇能力,这对推动“碳达峰”和“碳中和”进程具有重要意义. ...
Substantial role of macroalgae in marine carbon sequestration
1
2016
... 滨海蓝碳增汇技术是指增加易于管理的滨海蓝碳生态系统中由海洋生物驱动的碳通量及碳储量的相关技术.目前滨海蓝碳增汇的主要途径是红树林、盐沼、海草床、海藻场生态系统的保护与修复.全球红树林每年的CO2封存量为(1.7±0.2)Mg/hm2,红树林面积约为1.5×107 hm2,占世界森林总面积的0.4%;然而,在过去30年间,全球红树林面积约减少了1.0×106 hm2,预计到2100年末,因红树林减少导致的CO2排放量为2.4×109~3.4×109 Mg[3].全球盐沼的碳储量为(334.4±3.5)Mg/hm2,盐沼面积约为5.5×106 hm2,占滨海蓝碳生态系统总面积的4%[4];2000—2019年,全球盐沼面积总损失量约为1.4×105 hm2,相当于新加坡面积的2倍,这导致每年约增加了1.6×107 Mg的CO2排放量[5].全球海草床储存的有机碳约达2.0×1010 Mg,海草床面积约为7.9×107 hm2,占全球海洋面积的2%;自20世纪以来,全球海草床的年平均损失率为1.5%,导致每年有1.1×107~2.3×107 Mg有机碳加速分解[6].大型海藻是海岸带的主要初级生产者,其净初级生产力为1.0×109~2.0×109 Mg/a,海藻场面积为2.8×108~4.3×108 hm2[7];在过去50年间,由于海洋污染、捕捞、物种入侵、水产养殖以及全球变暖等原因,约60%的海藻场已退化[8].滨海蓝碳生态系统的退化显著削弱了其碳汇能力,因此,亟须修复滨海蓝碳生态系统,提高其碳汇能力,这对推动“碳达峰”和“碳中和”进程具有重要意义. ...
Potential role of seaweeds in climate change mitigation
1
2023
... 滨海蓝碳增汇技术是指增加易于管理的滨海蓝碳生态系统中由海洋生物驱动的碳通量及碳储量的相关技术.目前滨海蓝碳增汇的主要途径是红树林、盐沼、海草床、海藻场生态系统的保护与修复.全球红树林每年的CO2封存量为(1.7±0.2)Mg/hm2,红树林面积约为1.5×107 hm2,占世界森林总面积的0.4%;然而,在过去30年间,全球红树林面积约减少了1.0×106 hm2,预计到2100年末,因红树林减少导致的CO2排放量为2.4×109~3.4×109 Mg[3].全球盐沼的碳储量为(334.4±3.5)Mg/hm2,盐沼面积约为5.5×106 hm2,占滨海蓝碳生态系统总面积的4%[4];2000—2019年,全球盐沼面积总损失量约为1.4×105 hm2,相当于新加坡面积的2倍,这导致每年约增加了1.6×107 Mg的CO2排放量[5].全球海草床储存的有机碳约达2.0×1010 Mg,海草床面积约为7.9×107 hm2,占全球海洋面积的2%;自20世纪以来,全球海草床的年平均损失率为1.5%,导致每年有1.1×107~2.3×107 Mg有机碳加速分解[6].大型海藻是海岸带的主要初级生产者,其净初级生产力为1.0×109~2.0×109 Mg/a,海藻场面积为2.8×108~4.3×108 hm2[7];在过去50年间,由于海洋污染、捕捞、物种入侵、水产养殖以及全球变暖等原因,约60%的海藻场已退化[8].滨海蓝碳生态系统的退化显著削弱了其碳汇能力,因此,亟须修复滨海蓝碳生态系统,提高其碳汇能力,这对推动“碳达峰”和“碳中和”进程具有重要意义. ...
Nutrient removal from Chinese coastal waters by large-scale seaweed aquaculture
1
2017
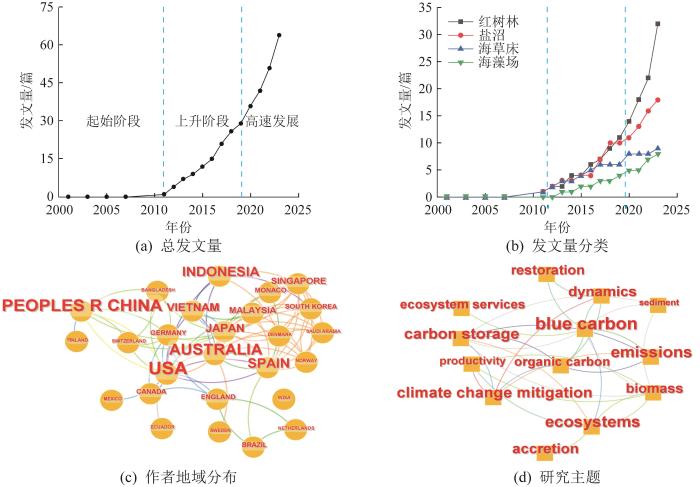
... 通过Web of Science网站检索到的关于滨海蓝碳增汇研究的发文情况如图2所示.由图2(a)可知,滨海蓝碳增汇的研究经历了起始阶段、上升阶段和高速发展阶段.2009年,联合国环境规划署(United Nations Environment Programme,UNEP)、联合国粮食及农业组织(Food and Agriculture Organization of the United Nations,FAO)和联合国教育科学文化组织政府间海洋学委员会(United Nations Educational, Scientific and Cultural Organization-Intergovernmental Oceanographic Commission,UNESCO-IOC)联合发布了《蓝色碳汇:健康海洋固碳作用的评估报告》(Blue Carbon: The Role of Healthy Oceans in Binding Carbon),首次提出“蓝碳”概念,即蓝碳指由海洋生物捕获的碳,滨海蓝碳增汇研究开始萌芽.随着气候变化共识的达成及2016年《巴黎协定》正式签署生效,滨海蓝碳增汇研究快速发展,相关发文量陡增.发文量分类如图2(b)所示.关于红树林蓝碳增汇的研究最多,盐沼和海草床次之,关于海藻场蓝碳增汇的研究最少.海藻场作为一种新兴的蓝碳生态系统,其碳汇功能和碳汇能力仍然存在一些争议,但随着越来越多的证据证明海藻场具有强大的碳汇潜力,其研究论文逐步增多.作者的地域分布如图2(c)所示.其中,美国、中国、澳大利亚、印度尼西亚的发文量较多.中国是世界上少数几个同时拥有红树林、盐沼、海草床三大蓝碳生态系统的国家之一,同时中国大型海藻产量占全球2/3左右[9];印度尼西亚拥有全世界最大面积的红树林.研究主题如图2(d)所示.研究主要集中在缓解气候变化、生态系统恢复及沉积物有机碳埋藏等. ...
Challenges and opportunities for achieving Sustainable Development Goals through restoration of Indonesia's mangroves
1
2023
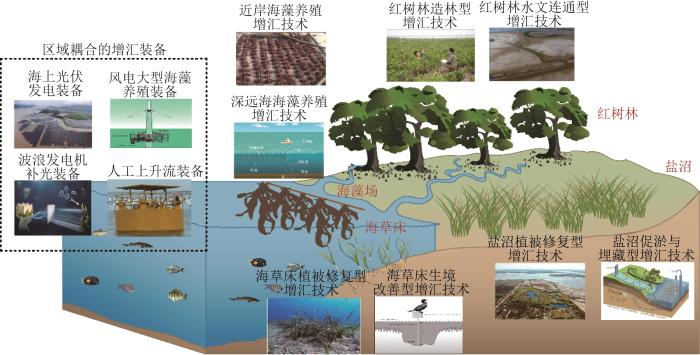
... 人口增长、粮食生产、工业和城市发展以及木材砍伐等因素导致红树林遭到大面积破坏,红树林造林型增汇技术则是通过植树造林恢复和扩大红树林面积,以此恢复和提高红树林的蓝碳储量,这是国内外广泛采用的主流的红树林增汇技术.例如:全球红树林品种最丰富的国家——印度尼西亚提出了“到2024年末红树林面积恢复6.0×105 hm2”的目标,该行动将使其全国土地CO2排放量减少16%[10]. ...
Replacing Spartina alterniflora with northward-afforested mangroves has the potential to acquire extra blue carbon
2
2024
... 红树林造林型增汇技术的广泛推广和成功应用,对全球红树林的分布格局、增汇格局产生了较大影响.例如:中国历史上红树林生态系统的最北端边界曾位于福建省福鼎市(27°20′ N)附近,1957年以来浙江省温州市(28°20′ N)成功引进红树林,实现了红树林北移.到2020年,浙江省的红树林面积达到386.8 hm2,北移3年后红树林的净碳吸收量达到1.1 Mg/(hm2·a)[11].Song等[12]预估在40年内红树林损失区域进行再造林后,可吸收6.7×108~6.9×108 Mg的CO2,并且相比于光滩造林,重新造林后的蓝碳储量约可增加60%. ...
... 红树林引进后北移(浙江温州)[11] ...
Mangrove reforestation provides greater blue carbon benefit than afforestation for mitigating global climate change
1
2023
... 红树林造林型增汇技术的广泛推广和成功应用,对全球红树林的分布格局、增汇格局产生了较大影响.例如:中国历史上红树林生态系统的最北端边界曾位于福建省福鼎市(27°20′ N)附近,1957年以来浙江省温州市(28°20′ N)成功引进红树林,实现了红树林北移.到2020年,浙江省的红树林面积达到386.8 hm2,北移3年后红树林的净碳吸收量达到1.1 Mg/(hm2·a)[11].Song等[12]预估在40年内红树林损失区域进行再造林后,可吸收6.7×108~6.9×108 Mg的CO2,并且相比于光滩造林,重新造林后的蓝碳储量约可增加60%. ...
Differences in ecosystem organic carbon stocks due to species selection and site elevation of restored mangrove forests
1
2023
... 红树林造林型增汇技术的关键包括造林位置和物种的选择等,环境因素对红树林的生长有着重要影响.从造林位置来看,随着种植场地海拔的降低,红树植物秋茄(Kandelia obovata)和白骨壤(Avicennia marina)的总生物量分别减少了48.8%和19.7%;从物种来看,白骨壤生态系统的有机碳储量高于秋茄生态系统的有机碳储量[13].因此,为了最大限度地提高红树林的固碳效果,可重点考虑在低海拔地区种植白骨壤.不过,红树林造林型增汇具有一定的局限性,例如其不适用于年平均降雨量少、淹水时间长、盐度高的区域. ...
海岸带蓝碳增汇: 理念、技术与未来建议
3
2023
... 填海造田、堤坝水利以及道路建造会阻断海洋与湿地之间的水文连通性,造成红树林退化.红树林水文连通型增汇通常需要修改或者移除水利工程以及直接连通水流,主要方式有降低防潮闸门的高度、拆除堤坝以及开通沟渠等.该增汇技术的原理主要包括:1)水文连通后,潮汐作用恢复,可以在淹水条件下营造厌氧环境,抑制有机质的分解[14];2)海水中丰富的硫酸盐沉积在湿地中,使得硫酸盐还原菌成为湿地中的优势菌群,这进一步缓解了由CH4排放造成的有机碳的损失[15];3)水文连通后排水功能恢复,使得水位下降且海水淹没时间缩短,红树林幼苗的根系或红树林种子在该水文条件下可以接触到沉积物,减少漂浮时间,保持其发芽和生长活力[16]. ...
... 泥沙的沉积速率和输入量是决定盐沼碳埋藏和碳储存能力的关键因素.盐沼促淤与埋藏型增汇技术主要通过提高潮汐作用下的泥沙输入量、增加盐沼沉积物的泥沙截留量、降低泥沙侵蚀量来维持盐沼的碳沉积与碳埋藏能力;恢复盐沼的水沙供给,促进生境的纵向堆积和横向延伸,扩展其向陆或向海的生存空间,提高碳沉积与碳埋藏的速率[14].主要技术措施有:1)引入临近水域的水沙或提高海岸的泥沙输入量,来提升盐沼的固沙能力.2)在潮流输入较弱的区域,在安全防护下将海堤向陆退缩,通过恢复海洋潮流的沉积作用塑造或改变盐沼生境类型,提高碳埋藏速率[25].例如:澳大利亚链谷湾的盐沼在沉降作用下经过5~10年转化为红树林湿地,沉积物堆积速率和碳埋藏速率分别提高了2倍和4倍[25].3)通过建造沙岛、阶地等来提高泥沙截留量,并减弱侵蚀效应[26].沙岛可通过降低水流速度和减弱海浪冲击促进泥沙悬浮,从而截留泥沙;阶地是由多条平行的沟坎组成的构筑物,它将湿地划分为多个小盆地,可促进泥沙沉积[14].4)将防洪堤、丁字坝等硬质堤坝改造为贻贝床、牡蛎礁等生物堤坝,可以促进泥沙供给,减弱海浪侵蚀[27].其中,牡蛎礁沉积物的泥沙含量高达60%,悬浮泥沙的质量浓度为50~370 mg/L[28]. ...
... [14].4)将防洪堤、丁字坝等硬质堤坝改造为贻贝床、牡蛎礁等生物堤坝,可以促进泥沙供给,减弱海浪侵蚀[27].其中,牡蛎礁沉积物的泥沙含量高达60%,悬浮泥沙的质量浓度为50~370 mg/L[28]. ...
海岸带蓝碳增汇: 理念、技术与未来建议
3
2023
... 填海造田、堤坝水利以及道路建造会阻断海洋与湿地之间的水文连通性,造成红树林退化.红树林水文连通型增汇通常需要修改或者移除水利工程以及直接连通水流,主要方式有降低防潮闸门的高度、拆除堤坝以及开通沟渠等.该增汇技术的原理主要包括:1)水文连通后,潮汐作用恢复,可以在淹水条件下营造厌氧环境,抑制有机质的分解[14];2)海水中丰富的硫酸盐沉积在湿地中,使得硫酸盐还原菌成为湿地中的优势菌群,这进一步缓解了由CH4排放造成的有机碳的损失[15];3)水文连通后排水功能恢复,使得水位下降且海水淹没时间缩短,红树林幼苗的根系或红树林种子在该水文条件下可以接触到沉积物,减少漂浮时间,保持其发芽和生长活力[16]. ...
... 泥沙的沉积速率和输入量是决定盐沼碳埋藏和碳储存能力的关键因素.盐沼促淤与埋藏型增汇技术主要通过提高潮汐作用下的泥沙输入量、增加盐沼沉积物的泥沙截留量、降低泥沙侵蚀量来维持盐沼的碳沉积与碳埋藏能力;恢复盐沼的水沙供给,促进生境的纵向堆积和横向延伸,扩展其向陆或向海的生存空间,提高碳沉积与碳埋藏的速率[14].主要技术措施有:1)引入临近水域的水沙或提高海岸的泥沙输入量,来提升盐沼的固沙能力.2)在潮流输入较弱的区域,在安全防护下将海堤向陆退缩,通过恢复海洋潮流的沉积作用塑造或改变盐沼生境类型,提高碳埋藏速率[25].例如:澳大利亚链谷湾的盐沼在沉降作用下经过5~10年转化为红树林湿地,沉积物堆积速率和碳埋藏速率分别提高了2倍和4倍[25].3)通过建造沙岛、阶地等来提高泥沙截留量,并减弱侵蚀效应[26].沙岛可通过降低水流速度和减弱海浪冲击促进泥沙悬浮,从而截留泥沙;阶地是由多条平行的沟坎组成的构筑物,它将湿地划分为多个小盆地,可促进泥沙沉积[14].4)将防洪堤、丁字坝等硬质堤坝改造为贻贝床、牡蛎礁等生物堤坝,可以促进泥沙供给,减弱海浪侵蚀[27].其中,牡蛎礁沉积物的泥沙含量高达60%,悬浮泥沙的质量浓度为50~370 mg/L[28]. ...
... [14].4)将防洪堤、丁字坝等硬质堤坝改造为贻贝床、牡蛎礁等生物堤坝,可以促进泥沙供给,减弱海浪侵蚀[27].其中,牡蛎礁沉积物的泥沙含量高达60%,悬浮泥沙的质量浓度为50~370 mg/L[28]. ...
Soils and sediments of coastal ecology: A global carbon sink
1
2021
... 填海造田、堤坝水利以及道路建造会阻断海洋与湿地之间的水文连通性,造成红树林退化.红树林水文连通型增汇通常需要修改或者移除水利工程以及直接连通水流,主要方式有降低防潮闸门的高度、拆除堤坝以及开通沟渠等.该增汇技术的原理主要包括:1)水文连通后,潮汐作用恢复,可以在淹水条件下营造厌氧环境,抑制有机质的分解[14];2)海水中丰富的硫酸盐沉积在湿地中,使得硫酸盐还原菌成为湿地中的优势菌群,这进一步缓解了由CH4排放造成的有机碳的损失[15];3)水文连通后排水功能恢复,使得水位下降且海水淹没时间缩短,红树林幼苗的根系或红树林种子在该水文条件下可以接触到沉积物,减少漂浮时间,保持其发芽和生长活力[16]. ...
Nurse species could facilitate the recruitment of mangrove seedlings after hydrological rehabilitation
1
2019
... 填海造田、堤坝水利以及道路建造会阻断海洋与湿地之间的水文连通性,造成红树林退化.红树林水文连通型增汇通常需要修改或者移除水利工程以及直接连通水流,主要方式有降低防潮闸门的高度、拆除堤坝以及开通沟渠等.该增汇技术的原理主要包括:1)水文连通后,潮汐作用恢复,可以在淹水条件下营造厌氧环境,抑制有机质的分解[14];2)海水中丰富的硫酸盐沉积在湿地中,使得硫酸盐还原菌成为湿地中的优势菌群,这进一步缓解了由CH4排放造成的有机碳的损失[15];3)水文连通后排水功能恢复,使得水位下降且海水淹没时间缩短,红树林幼苗的根系或红树林种子在该水文条件下可以接触到沉积物,减少漂浮时间,保持其发芽和生长活力[16]. ...
High greenhouse gas emissions mitigation benefits from mangrove rehabilitation in Sulawesi, Indonesia
2
2019
... 目前,国内外已有较多红树林水文连通型增汇技术的工程应用实例.例如:印度尼西亚苏拉威西岛的废弃池塘恢复水文活动后,促进了红树林再生,该修复区域每年大约减少了27.6 Mg/hm2的CO2排放 [17].不过,由于后续河流携带的泥沙不断沉淀,主要的渠道容易堵塞,需要经常疏浚,因此增汇管护时在水文修复方面需投入的资金较多. ...
... 威西岛)[17] ...
Revitalizing coastal saline-alkali soil with biochar application for improved crop growth
2
2022
... 受到外来物种入侵、海岸工程建设和海平面上升等因素的影响,盐沼本土植物会大量死亡甚至全部消失.盐沼植被修复型增汇技术主要通过促进盐沼植被恢复和生长来提升由植物主导的内源碳库的固碳功能.其中,生物炭和微生物菌剂具有提供养分(氮、磷和钾等)和改良土壤(优化pH值、阳离子交换能力、盐度、容重及减少污染物)等功能,施用后有利于植物根系发育,促进植物生长,并从大气中吸收更多的CO2,因此是值得关注的盐沼植被修复新材料.例如:为期2年的原位实验表明,添加生物炭后,盐沼土壤有机质增加了48.5%~139.6%,植物生长量提高了49.2%~56.7%[18];通过60 d的盆栽实验发现,施加1%的互花米草生物炭后,碱蓬幼苗的生物量约提高了100%[19];Sood等[20]发现,施加木霉菌(Trichderma)后,叶片中的钾浓度升高,植物根系吸收养分的能力提高;施加44.9 kg /hm2的木霉菌后,盐沼植物海三棱藨草的植株增长率可达669%,土壤的脲酶活性和碱性磷酸酶活性显著提高[21].此外,生物炭和微生物菌剂在缓解植物受到化感胁迫方面也具有较大的潜力.外来入侵生物互花米草会分泌化感物质如庚烷、棕榈酸、硬脂酸、2, 4-二叔丁基酚和氢化肉桂酸等[22],这些化感物质通过干扰光合作用、阻断三磷酸腺苷的合成和破坏植物的正常代谢来抑制周围盐沼本土植物的生长,即便清除了互花米草,残留的化感物质在短期内也无法完全降解消除,而添加生物炭和微生物菌剂可以有效缓解这些化感物质对盐沼本土植物的胁迫作用.例如:磷酸改性后的生物炭可以去除40%以上的酚酸类化感物质[23],内生真菌如枫香拟茎点霉(Phomopsis liquidambari)可以有效降解4-羟基苯甲酸和芥子酸[24]. ...
... (江苏盐城)[18] ...
Biochar-amended coastal wetland soil enhances growth of Suaeda salsa and alters rhizosphere soil nutrients and microbial communities
1
2021
... 受到外来物种入侵、海岸工程建设和海平面上升等因素的影响,盐沼本土植物会大量死亡甚至全部消失.盐沼植被修复型增汇技术主要通过促进盐沼植被恢复和生长来提升由植物主导的内源碳库的固碳功能.其中,生物炭和微生物菌剂具有提供养分(氮、磷和钾等)和改良土壤(优化pH值、阳离子交换能力、盐度、容重及减少污染物)等功能,施用后有利于植物根系发育,促进植物生长,并从大气中吸收更多的CO2,因此是值得关注的盐沼植被修复新材料.例如:为期2年的原位实验表明,添加生物炭后,盐沼土壤有机质增加了48.5%~139.6%,植物生长量提高了49.2%~56.7%[18];通过60 d的盆栽实验发现,施加1%的互花米草生物炭后,碱蓬幼苗的生物量约提高了100%[19];Sood等[20]发现,施加木霉菌(Trichderma)后,叶片中的钾浓度升高,植物根系吸收养分的能力提高;施加44.9 kg /hm2的木霉菌后,盐沼植物海三棱藨草的植株增长率可达669%,土壤的脲酶活性和碱性磷酸酶活性显著提高[21].此外,生物炭和微生物菌剂在缓解植物受到化感胁迫方面也具有较大的潜力.外来入侵生物互花米草会分泌化感物质如庚烷、棕榈酸、硬脂酸、2, 4-二叔丁基酚和氢化肉桂酸等[22],这些化感物质通过干扰光合作用、阻断三磷酸腺苷的合成和破坏植物的正常代谢来抑制周围盐沼本土植物的生长,即便清除了互花米草,残留的化感物质在短期内也无法完全降解消除,而添加生物炭和微生物菌剂可以有效缓解这些化感物质对盐沼本土植物的胁迫作用.例如:磷酸改性后的生物炭可以去除40%以上的酚酸类化感物质[23],内生真菌如枫香拟茎点霉(Phomopsis liquidambari)可以有效降解4-羟基苯甲酸和芥子酸[24]. ...
Trichoderma: the secrets of a multitalented biocontrol agent
1
2020
... 受到外来物种入侵、海岸工程建设和海平面上升等因素的影响,盐沼本土植物会大量死亡甚至全部消失.盐沼植被修复型增汇技术主要通过促进盐沼植被恢复和生长来提升由植物主导的内源碳库的固碳功能.其中,生物炭和微生物菌剂具有提供养分(氮、磷和钾等)和改良土壤(优化pH值、阳离子交换能力、盐度、容重及减少污染物)等功能,施用后有利于植物根系发育,促进植物生长,并从大气中吸收更多的CO2,因此是值得关注的盐沼植被修复新材料.例如:为期2年的原位实验表明,添加生物炭后,盐沼土壤有机质增加了48.5%~139.6%,植物生长量提高了49.2%~56.7%[18];通过60 d的盆栽实验发现,施加1%的互花米草生物炭后,碱蓬幼苗的生物量约提高了100%[19];Sood等[20]发现,施加木霉菌(Trichderma)后,叶片中的钾浓度升高,植物根系吸收养分的能力提高;施加44.9 kg /hm2的木霉菌后,盐沼植物海三棱藨草的植株增长率可达669%,土壤的脲酶活性和碱性磷酸酶活性显著提高[21].此外,生物炭和微生物菌剂在缓解植物受到化感胁迫方面也具有较大的潜力.外来入侵生物互花米草会分泌化感物质如庚烷、棕榈酸、硬脂酸、2, 4-二叔丁基酚和氢化肉桂酸等[22],这些化感物质通过干扰光合作用、阻断三磷酸腺苷的合成和破坏植物的正常代谢来抑制周围盐沼本土植物的生长,即便清除了互花米草,残留的化感物质在短期内也无法完全降解消除,而添加生物炭和微生物菌剂可以有效缓解这些化感物质对盐沼本土植物的胁迫作用.例如:磷酸改性后的生物炭可以去除40%以上的酚酸类化感物质[23],内生真菌如枫香拟茎点霉(Phomopsis liquidambari)可以有效降解4-羟基苯甲酸和芥子酸[24]. ...
木霉菌和复合微生物制剂在海三棱藨草湿地修复中的应用研究
1
2024
... 受到外来物种入侵、海岸工程建设和海平面上升等因素的影响,盐沼本土植物会大量死亡甚至全部消失.盐沼植被修复型增汇技术主要通过促进盐沼植被恢复和生长来提升由植物主导的内源碳库的固碳功能.其中,生物炭和微生物菌剂具有提供养分(氮、磷和钾等)和改良土壤(优化pH值、阳离子交换能力、盐度、容重及减少污染物)等功能,施用后有利于植物根系发育,促进植物生长,并从大气中吸收更多的CO2,因此是值得关注的盐沼植被修复新材料.例如:为期2年的原位实验表明,添加生物炭后,盐沼土壤有机质增加了48.5%~139.6%,植物生长量提高了49.2%~56.7%[18];通过60 d的盆栽实验发现,施加1%的互花米草生物炭后,碱蓬幼苗的生物量约提高了100%[19];Sood等[20]发现,施加木霉菌(Trichderma)后,叶片中的钾浓度升高,植物根系吸收养分的能力提高;施加44.9 kg /hm2的木霉菌后,盐沼植物海三棱藨草的植株增长率可达669%,土壤的脲酶活性和碱性磷酸酶活性显著提高[21].此外,生物炭和微生物菌剂在缓解植物受到化感胁迫方面也具有较大的潜力.外来入侵生物互花米草会分泌化感物质如庚烷、棕榈酸、硬脂酸、2, 4-二叔丁基酚和氢化肉桂酸等[22],这些化感物质通过干扰光合作用、阻断三磷酸腺苷的合成和破坏植物的正常代谢来抑制周围盐沼本土植物的生长,即便清除了互花米草,残留的化感物质在短期内也无法完全降解消除,而添加生物炭和微生物菌剂可以有效缓解这些化感物质对盐沼本土植物的胁迫作用.例如:磷酸改性后的生物炭可以去除40%以上的酚酸类化感物质[23],内生真菌如枫香拟茎点霉(Phomopsis liquidambari)可以有效降解4-羟基苯甲酸和芥子酸[24]. ...
木霉菌和复合微生物制剂在海三棱藨草湿地修复中的应用研究
1
2024
... 受到外来物种入侵、海岸工程建设和海平面上升等因素的影响,盐沼本土植物会大量死亡甚至全部消失.盐沼植被修复型增汇技术主要通过促进盐沼植被恢复和生长来提升由植物主导的内源碳库的固碳功能.其中,生物炭和微生物菌剂具有提供养分(氮、磷和钾等)和改良土壤(优化pH值、阳离子交换能力、盐度、容重及减少污染物)等功能,施用后有利于植物根系发育,促进植物生长,并从大气中吸收更多的CO2,因此是值得关注的盐沼植被修复新材料.例如:为期2年的原位实验表明,添加生物炭后,盐沼土壤有机质增加了48.5%~139.6%,植物生长量提高了49.2%~56.7%[18];通过60 d的盆栽实验发现,施加1%的互花米草生物炭后,碱蓬幼苗的生物量约提高了100%[19];Sood等[20]发现,施加木霉菌(Trichderma)后,叶片中的钾浓度升高,植物根系吸收养分的能力提高;施加44.9 kg /hm2的木霉菌后,盐沼植物海三棱藨草的植株增长率可达669%,土壤的脲酶活性和碱性磷酸酶活性显著提高[21].此外,生物炭和微生物菌剂在缓解植物受到化感胁迫方面也具有较大的潜力.外来入侵生物互花米草会分泌化感物质如庚烷、棕榈酸、硬脂酸、2, 4-二叔丁基酚和氢化肉桂酸等[22],这些化感物质通过干扰光合作用、阻断三磷酸腺苷的合成和破坏植物的正常代谢来抑制周围盐沼本土植物的生长,即便清除了互花米草,残留的化感物质在短期内也无法完全降解消除,而添加生物炭和微生物菌剂可以有效缓解这些化感物质对盐沼本土植物的胁迫作用.例如:磷酸改性后的生物炭可以去除40%以上的酚酸类化感物质[23],内生真菌如枫香拟茎点霉(Phomopsis liquidambari)可以有效降解4-羟基苯甲酸和芥子酸[24]. ...
Biochar mitigates allelopathy through regulating allelochemical generation from plants and accumulation in soil
1
2022
... 受到外来物种入侵、海岸工程建设和海平面上升等因素的影响,盐沼本土植物会大量死亡甚至全部消失.盐沼植被修复型增汇技术主要通过促进盐沼植被恢复和生长来提升由植物主导的内源碳库的固碳功能.其中,生物炭和微生物菌剂具有提供养分(氮、磷和钾等)和改良土壤(优化pH值、阳离子交换能力、盐度、容重及减少污染物)等功能,施用后有利于植物根系发育,促进植物生长,并从大气中吸收更多的CO2,因此是值得关注的盐沼植被修复新材料.例如:为期2年的原位实验表明,添加生物炭后,盐沼土壤有机质增加了48.5%~139.6%,植物生长量提高了49.2%~56.7%[18];通过60 d的盆栽实验发现,施加1%的互花米草生物炭后,碱蓬幼苗的生物量约提高了100%[19];Sood等[20]发现,施加木霉菌(Trichderma)后,叶片中的钾浓度升高,植物根系吸收养分的能力提高;施加44.9 kg /hm2的木霉菌后,盐沼植物海三棱藨草的植株增长率可达669%,土壤的脲酶活性和碱性磷酸酶活性显著提高[21].此外,生物炭和微生物菌剂在缓解植物受到化感胁迫方面也具有较大的潜力.外来入侵生物互花米草会分泌化感物质如庚烷、棕榈酸、硬脂酸、2, 4-二叔丁基酚和氢化肉桂酸等[22],这些化感物质通过干扰光合作用、阻断三磷酸腺苷的合成和破坏植物的正常代谢来抑制周围盐沼本土植物的生长,即便清除了互花米草,残留的化感物质在短期内也无法完全降解消除,而添加生物炭和微生物菌剂可以有效缓解这些化感物质对盐沼本土植物的胁迫作用.例如:磷酸改性后的生物炭可以去除40%以上的酚酸类化感物质[23],内生真菌如枫香拟茎点霉(Phomopsis liquidambari)可以有效降解4-羟基苯甲酸和芥子酸[24]. ...
Inhibition of continuous cropping obstacle of celery by chemically modified biochar: An efficient approach to decrease bioavailability of phenolic allelochemicals
1
2023
... 受到外来物种入侵、海岸工程建设和海平面上升等因素的影响,盐沼本土植物会大量死亡甚至全部消失.盐沼植被修复型增汇技术主要通过促进盐沼植被恢复和生长来提升由植物主导的内源碳库的固碳功能.其中,生物炭和微生物菌剂具有提供养分(氮、磷和钾等)和改良土壤(优化pH值、阳离子交换能力、盐度、容重及减少污染物)等功能,施用后有利于植物根系发育,促进植物生长,并从大气中吸收更多的CO2,因此是值得关注的盐沼植被修复新材料.例如:为期2年的原位实验表明,添加生物炭后,盐沼土壤有机质增加了48.5%~139.6%,植物生长量提高了49.2%~56.7%[18];通过60 d的盆栽实验发现,施加1%的互花米草生物炭后,碱蓬幼苗的生物量约提高了100%[19];Sood等[20]发现,施加木霉菌(Trichderma)后,叶片中的钾浓度升高,植物根系吸收养分的能力提高;施加44.9 kg /hm2的木霉菌后,盐沼植物海三棱藨草的植株增长率可达669%,土壤的脲酶活性和碱性磷酸酶活性显著提高[21].此外,生物炭和微生物菌剂在缓解植物受到化感胁迫方面也具有较大的潜力.外来入侵生物互花米草会分泌化感物质如庚烷、棕榈酸、硬脂酸、2, 4-二叔丁基酚和氢化肉桂酸等[22],这些化感物质通过干扰光合作用、阻断三磷酸腺苷的合成和破坏植物的正常代谢来抑制周围盐沼本土植物的生长,即便清除了互花米草,残留的化感物质在短期内也无法完全降解消除,而添加生物炭和微生物菌剂可以有效缓解这些化感物质对盐沼本土植物的胁迫作用.例如:磷酸改性后的生物炭可以去除40%以上的酚酸类化感物质[23],内生真菌如枫香拟茎点霉(Phomopsis liquidambari)可以有效降解4-羟基苯甲酸和芥子酸[24]. ...
Biodegradation of 4-hydroxybenzoic acid by Phomopsis liquidambari
1
2011
... 受到外来物种入侵、海岸工程建设和海平面上升等因素的影响,盐沼本土植物会大量死亡甚至全部消失.盐沼植被修复型增汇技术主要通过促进盐沼植被恢复和生长来提升由植物主导的内源碳库的固碳功能.其中,生物炭和微生物菌剂具有提供养分(氮、磷和钾等)和改良土壤(优化pH值、阳离子交换能力、盐度、容重及减少污染物)等功能,施用后有利于植物根系发育,促进植物生长,并从大气中吸收更多的CO2,因此是值得关注的盐沼植被修复新材料.例如:为期2年的原位实验表明,添加生物炭后,盐沼土壤有机质增加了48.5%~139.6%,植物生长量提高了49.2%~56.7%[18];通过60 d的盆栽实验发现,施加1%的互花米草生物炭后,碱蓬幼苗的生物量约提高了100%[19];Sood等[20]发现,施加木霉菌(Trichderma)后,叶片中的钾浓度升高,植物根系吸收养分的能力提高;施加44.9 kg /hm2的木霉菌后,盐沼植物海三棱藨草的植株增长率可达669%,土壤的脲酶活性和碱性磷酸酶活性显著提高[21].此外,生物炭和微生物菌剂在缓解植物受到化感胁迫方面也具有较大的潜力.外来入侵生物互花米草会分泌化感物质如庚烷、棕榈酸、硬脂酸、2, 4-二叔丁基酚和氢化肉桂酸等[22],这些化感物质通过干扰光合作用、阻断三磷酸腺苷的合成和破坏植物的正常代谢来抑制周围盐沼本土植物的生长,即便清除了互花米草,残留的化感物质在短期内也无法完全降解消除,而添加生物炭和微生物菌剂可以有效缓解这些化感物质对盐沼本土植物的胁迫作用.例如:磷酸改性后的生物炭可以去除40%以上的酚酸类化感物质[23],内生真菌如枫香拟茎点霉(Phomopsis liquidambari)可以有效降解4-羟基苯甲酸和芥子酸[24]. ...
Wetland carbon storage controlled by millennial-scale variation in relative sea-level rise
3
2019
... 泥沙的沉积速率和输入量是决定盐沼碳埋藏和碳储存能力的关键因素.盐沼促淤与埋藏型增汇技术主要通过提高潮汐作用下的泥沙输入量、增加盐沼沉积物的泥沙截留量、降低泥沙侵蚀量来维持盐沼的碳沉积与碳埋藏能力;恢复盐沼的水沙供给,促进生境的纵向堆积和横向延伸,扩展其向陆或向海的生存空间,提高碳沉积与碳埋藏的速率[14].主要技术措施有:1)引入临近水域的水沙或提高海岸的泥沙输入量,来提升盐沼的固沙能力.2)在潮流输入较弱的区域,在安全防护下将海堤向陆退缩,通过恢复海洋潮流的沉积作用塑造或改变盐沼生境类型,提高碳埋藏速率[25].例如:澳大利亚链谷湾的盐沼在沉降作用下经过5~10年转化为红树林湿地,沉积物堆积速率和碳埋藏速率分别提高了2倍和4倍[25].3)通过建造沙岛、阶地等来提高泥沙截留量,并减弱侵蚀效应[26].沙岛可通过降低水流速度和减弱海浪冲击促进泥沙悬浮,从而截留泥沙;阶地是由多条平行的沟坎组成的构筑物,它将湿地划分为多个小盆地,可促进泥沙沉积[14].4)将防洪堤、丁字坝等硬质堤坝改造为贻贝床、牡蛎礁等生物堤坝,可以促进泥沙供给,减弱海浪侵蚀[27].其中,牡蛎礁沉积物的泥沙含量高达60%,悬浮泥沙的质量浓度为50~370 mg/L[28]. ...
... [25].3)通过建造沙岛、阶地等来提高泥沙截留量,并减弱侵蚀效应[26].沙岛可通过降低水流速度和减弱海浪冲击促进泥沙悬浮,从而截留泥沙;阶地是由多条平行的沟坎组成的构筑物,它将湿地划分为多个小盆地,可促进泥沙沉积[14].4)将防洪堤、丁字坝等硬质堤坝改造为贻贝床、牡蛎礁等生物堤坝,可以促进泥沙供给,减弱海浪侵蚀[27].其中,牡蛎礁沉积物的泥沙含量高达60%,悬浮泥沙的质量浓度为50~370 mg/L[28]. ...
... 链谷湾)[25] ...
A review of sediment diversion in the Mississippi River Deltaic Plain
1
2019
... 泥沙的沉积速率和输入量是决定盐沼碳埋藏和碳储存能力的关键因素.盐沼促淤与埋藏型增汇技术主要通过提高潮汐作用下的泥沙输入量、增加盐沼沉积物的泥沙截留量、降低泥沙侵蚀量来维持盐沼的碳沉积与碳埋藏能力;恢复盐沼的水沙供给,促进生境的纵向堆积和横向延伸,扩展其向陆或向海的生存空间,提高碳沉积与碳埋藏的速率[14].主要技术措施有:1)引入临近水域的水沙或提高海岸的泥沙输入量,来提升盐沼的固沙能力.2)在潮流输入较弱的区域,在安全防护下将海堤向陆退缩,通过恢复海洋潮流的沉积作用塑造或改变盐沼生境类型,提高碳埋藏速率[25].例如:澳大利亚链谷湾的盐沼在沉降作用下经过5~10年转化为红树林湿地,沉积物堆积速率和碳埋藏速率分别提高了2倍和4倍[25].3)通过建造沙岛、阶地等来提高泥沙截留量,并减弱侵蚀效应[26].沙岛可通过降低水流速度和减弱海浪冲击促进泥沙悬浮,从而截留泥沙;阶地是由多条平行的沟坎组成的构筑物,它将湿地划分为多个小盆地,可促进泥沙沉积[14].4)将防洪堤、丁字坝等硬质堤坝改造为贻贝床、牡蛎礁等生物堤坝,可以促进泥沙供给,减弱海浪侵蚀[27].其中,牡蛎礁沉积物的泥沙含量高达60%,悬浮泥沙的质量浓度为50~370 mg/L[28]. ...
Understanding the conditionality of ecosystem services: The effect of tidal flat morphology and oyster reef characteristics on sediment stabilization by oyster reefs
1
2018
... 泥沙的沉积速率和输入量是决定盐沼碳埋藏和碳储存能力的关键因素.盐沼促淤与埋藏型增汇技术主要通过提高潮汐作用下的泥沙输入量、增加盐沼沉积物的泥沙截留量、降低泥沙侵蚀量来维持盐沼的碳沉积与碳埋藏能力;恢复盐沼的水沙供给,促进生境的纵向堆积和横向延伸,扩展其向陆或向海的生存空间,提高碳沉积与碳埋藏的速率[14].主要技术措施有:1)引入临近水域的水沙或提高海岸的泥沙输入量,来提升盐沼的固沙能力.2)在潮流输入较弱的区域,在安全防护下将海堤向陆退缩,通过恢复海洋潮流的沉积作用塑造或改变盐沼生境类型,提高碳埋藏速率[25].例如:澳大利亚链谷湾的盐沼在沉降作用下经过5~10年转化为红树林湿地,沉积物堆积速率和碳埋藏速率分别提高了2倍和4倍[25].3)通过建造沙岛、阶地等来提高泥沙截留量,并减弱侵蚀效应[26].沙岛可通过降低水流速度和减弱海浪冲击促进泥沙悬浮,从而截留泥沙;阶地是由多条平行的沟坎组成的构筑物,它将湿地划分为多个小盆地,可促进泥沙沉积[14].4)将防洪堤、丁字坝等硬质堤坝改造为贻贝床、牡蛎礁等生物堤坝,可以促进泥沙供给,减弱海浪侵蚀[27].其中,牡蛎礁沉积物的泥沙含量高达60%,悬浮泥沙的质量浓度为50~370 mg/L[28]. ...
Geomorphology and sediment dynamics of the Liyashan oyster reefs, Jiangsu Coast, China
1
2021
... 泥沙的沉积速率和输入量是决定盐沼碳埋藏和碳储存能力的关键因素.盐沼促淤与埋藏型增汇技术主要通过提高潮汐作用下的泥沙输入量、增加盐沼沉积物的泥沙截留量、降低泥沙侵蚀量来维持盐沼的碳沉积与碳埋藏能力;恢复盐沼的水沙供给,促进生境的纵向堆积和横向延伸,扩展其向陆或向海的生存空间,提高碳沉积与碳埋藏的速率[14].主要技术措施有:1)引入临近水域的水沙或提高海岸的泥沙输入量,来提升盐沼的固沙能力.2)在潮流输入较弱的区域,在安全防护下将海堤向陆退缩,通过恢复海洋潮流的沉积作用塑造或改变盐沼生境类型,提高碳埋藏速率[25].例如:澳大利亚链谷湾的盐沼在沉降作用下经过5~10年转化为红树林湿地,沉积物堆积速率和碳埋藏速率分别提高了2倍和4倍[25].3)通过建造沙岛、阶地等来提高泥沙截留量,并减弱侵蚀效应[26].沙岛可通过降低水流速度和减弱海浪冲击促进泥沙悬浮,从而截留泥沙;阶地是由多条平行的沟坎组成的构筑物,它将湿地划分为多个小盆地,可促进泥沙沉积[14].4)将防洪堤、丁字坝等硬质堤坝改造为贻贝床、牡蛎礁等生物堤坝,可以促进泥沙供给,减弱海浪侵蚀[27].其中,牡蛎礁沉积物的泥沙含量高达60%,悬浮泥沙的质量浓度为50~370 mg/L[28]. ...
Germination and early-stage development in the seagrass, Zostera muelleri Irmisch ex Asch in response to multiple stressors
1
2016
... 受到水产养殖、船舶运输和有害藻华的影响,海草大面积减少,导致裸露的沉积物中封存的有机碳加速分解.海草床植被修复型增汇技术主要通过提高海草种子萌芽率和海草移植成功率的方式来恢复海草床的碳封存能力.在提高海草种子萌芽率方面,已有多种因地制宜的技术被成功研发.例如:用淡水浸泡种子或在种子表面制造划痕,来有效诱导海草种子发芽[29];甲壳动物对海草种子的捕食会降低海草床的恢复成功率,因此将海菖蒲(Enhalus acoroides)种子装入网袋且使其埋藏深度为2 cm,以避免生物捕食造成的损失,种子发芽率可高达96%[30].在提高海草移植成功率方面,修复海草床植被的常用方法是草皮法、草块法和根状茎法.为了提高海草移植效率,澳大利亚研发了移植海草的专用机械设备ECOSUB1和ECOSUB2[31-32],提高了海草移植的机械化水平.另外,海草移植后容易被海浪冲走,因此还需对其进行固定.目前常用的海草移植固定方法有订书钉法、框架法和贝壳法.研究发现[33],采用订书钉法把海草两端固定于潮滩底部,4个月后海草的覆盖率可达到57.8%.被修复的海草床的固碳能力与天然海草床相近.例如:在澳大利亚西澳大利亚州牡蛎港的海岸移植海草18年后,海草床的碳埋藏率达到了0.3 Mg/(hm2·a)[34]. ...
基于种子法的海菖蒲海草床恢复
1
2019
... 受到水产养殖、船舶运输和有害藻华的影响,海草大面积减少,导致裸露的沉积物中封存的有机碳加速分解.海草床植被修复型增汇技术主要通过提高海草种子萌芽率和海草移植成功率的方式来恢复海草床的碳封存能力.在提高海草种子萌芽率方面,已有多种因地制宜的技术被成功研发.例如:用淡水浸泡种子或在种子表面制造划痕,来有效诱导海草种子发芽[29];甲壳动物对海草种子的捕食会降低海草床的恢复成功率,因此将海菖蒲(Enhalus acoroides)种子装入网袋且使其埋藏深度为2 cm,以避免生物捕食造成的损失,种子发芽率可高达96%[30].在提高海草移植成功率方面,修复海草床植被的常用方法是草皮法、草块法和根状茎法.为了提高海草移植效率,澳大利亚研发了移植海草的专用机械设备ECOSUB1和ECOSUB2[31-32],提高了海草移植的机械化水平.另外,海草移植后容易被海浪冲走,因此还需对其进行固定.目前常用的海草移植固定方法有订书钉法、框架法和贝壳法.研究发现[33],采用订书钉法把海草两端固定于潮滩底部,4个月后海草的覆盖率可达到57.8%.被修复的海草床的固碳能力与天然海草床相近.例如:在澳大利亚西澳大利亚州牡蛎港的海岸移植海草18年后,海草床的碳埋藏率达到了0.3 Mg/(hm2·a)[34]. ...
基于种子法的海菖蒲海草床恢复
1
2019
... 受到水产养殖、船舶运输和有害藻华的影响,海草大面积减少,导致裸露的沉积物中封存的有机碳加速分解.海草床植被修复型增汇技术主要通过提高海草种子萌芽率和海草移植成功率的方式来恢复海草床的碳封存能力.在提高海草种子萌芽率方面,已有多种因地制宜的技术被成功研发.例如:用淡水浸泡种子或在种子表面制造划痕,来有效诱导海草种子发芽[29];甲壳动物对海草种子的捕食会降低海草床的恢复成功率,因此将海菖蒲(Enhalus acoroides)种子装入网袋且使其埋藏深度为2 cm,以避免生物捕食造成的损失,种子发芽率可高达96%[30].在提高海草移植成功率方面,修复海草床植被的常用方法是草皮法、草块法和根状茎法.为了提高海草移植效率,澳大利亚研发了移植海草的专用机械设备ECOSUB1和ECOSUB2[31-32],提高了海草移植的机械化水平.另外,海草移植后容易被海浪冲走,因此还需对其进行固定.目前常用的海草移植固定方法有订书钉法、框架法和贝壳法.研究发现[33],采用订书钉法把海草两端固定于潮滩底部,4个月后海草的覆盖率可达到57.8%.被修复的海草床的固碳能力与天然海草床相近.例如:在澳大利亚西澳大利亚州牡蛎港的海岸移植海草18年后,海草床的碳埋藏率达到了0.3 Mg/(hm2·a)[34]. ...
Mechanical seagrass transplantation in Western Australia
1
2001
... 受到水产养殖、船舶运输和有害藻华的影响,海草大面积减少,导致裸露的沉积物中封存的有机碳加速分解.海草床植被修复型增汇技术主要通过提高海草种子萌芽率和海草移植成功率的方式来恢复海草床的碳封存能力.在提高海草种子萌芽率方面,已有多种因地制宜的技术被成功研发.例如:用淡水浸泡种子或在种子表面制造划痕,来有效诱导海草种子发芽[29];甲壳动物对海草种子的捕食会降低海草床的恢复成功率,因此将海菖蒲(Enhalus acoroides)种子装入网袋且使其埋藏深度为2 cm,以避免生物捕食造成的损失,种子发芽率可高达96%[30].在提高海草移植成功率方面,修复海草床植被的常用方法是草皮法、草块法和根状茎法.为了提高海草移植效率,澳大利亚研发了移植海草的专用机械设备ECOSUB1和ECOSUB2[31-32],提高了海草移植的机械化水平.另外,海草移植后容易被海浪冲走,因此还需对其进行固定.目前常用的海草移植固定方法有订书钉法、框架法和贝壳法.研究发现[33],采用订书钉法把海草两端固定于潮滩底部,4个月后海草的覆盖率可达到57.8%.被修复的海草床的固碳能力与天然海草床相近.例如:在澳大利亚西澳大利亚州牡蛎港的海岸移植海草18年后,海草床的碳埋藏率达到了0.3 Mg/(hm2·a)[34]. ...
Improving mechanical seagrass transplantation
1
2001
... 受到水产养殖、船舶运输和有害藻华的影响,海草大面积减少,导致裸露的沉积物中封存的有机碳加速分解.海草床植被修复型增汇技术主要通过提高海草种子萌芽率和海草移植成功率的方式来恢复海草床的碳封存能力.在提高海草种子萌芽率方面,已有多种因地制宜的技术被成功研发.例如:用淡水浸泡种子或在种子表面制造划痕,来有效诱导海草种子发芽[29];甲壳动物对海草种子的捕食会降低海草床的恢复成功率,因此将海菖蒲(Enhalus acoroides)种子装入网袋且使其埋藏深度为2 cm,以避免生物捕食造成的损失,种子发芽率可高达96%[30].在提高海草移植成功率方面,修复海草床植被的常用方法是草皮法、草块法和根状茎法.为了提高海草移植效率,澳大利亚研发了移植海草的专用机械设备ECOSUB1和ECOSUB2[31-32],提高了海草移植的机械化水平.另外,海草移植后容易被海浪冲走,因此还需对其进行固定.目前常用的海草移植固定方法有订书钉法、框架法和贝壳法.研究发现[33],采用订书钉法把海草两端固定于潮滩底部,4个月后海草的覆盖率可达到57.8%.被修复的海草床的固碳能力与天然海草床相近.例如:在澳大利亚西澳大利亚州牡蛎港的海岸移植海草18年后,海草床的碳埋藏率达到了0.3 Mg/(hm2·a)[34]. ...
移植操作胁迫对天鹅湖大叶藻存活、生长及光合色素含量的影响
1
2014
... 受到水产养殖、船舶运输和有害藻华的影响,海草大面积减少,导致裸露的沉积物中封存的有机碳加速分解.海草床植被修复型增汇技术主要通过提高海草种子萌芽率和海草移植成功率的方式来恢复海草床的碳封存能力.在提高海草种子萌芽率方面,已有多种因地制宜的技术被成功研发.例如:用淡水浸泡种子或在种子表面制造划痕,来有效诱导海草种子发芽[29];甲壳动物对海草种子的捕食会降低海草床的恢复成功率,因此将海菖蒲(Enhalus acoroides)种子装入网袋且使其埋藏深度为2 cm,以避免生物捕食造成的损失,种子发芽率可高达96%[30].在提高海草移植成功率方面,修复海草床植被的常用方法是草皮法、草块法和根状茎法.为了提高海草移植效率,澳大利亚研发了移植海草的专用机械设备ECOSUB1和ECOSUB2[31-32],提高了海草移植的机械化水平.另外,海草移植后容易被海浪冲走,因此还需对其进行固定.目前常用的海草移植固定方法有订书钉法、框架法和贝壳法.研究发现[33],采用订书钉法把海草两端固定于潮滩底部,4个月后海草的覆盖率可达到57.8%.被修复的海草床的固碳能力与天然海草床相近.例如:在澳大利亚西澳大利亚州牡蛎港的海岸移植海草18年后,海草床的碳埋藏率达到了0.3 Mg/(hm2·a)[34]. ...
移植操作胁迫对天鹅湖大叶藻存活、生长及光合色素含量的影响
1
2014
... 受到水产养殖、船舶运输和有害藻华的影响,海草大面积减少,导致裸露的沉积物中封存的有机碳加速分解.海草床植被修复型增汇技术主要通过提高海草种子萌芽率和海草移植成功率的方式来恢复海草床的碳封存能力.在提高海草种子萌芽率方面,已有多种因地制宜的技术被成功研发.例如:用淡水浸泡种子或在种子表面制造划痕,来有效诱导海草种子发芽[29];甲壳动物对海草种子的捕食会降低海草床的恢复成功率,因此将海菖蒲(Enhalus acoroides)种子装入网袋且使其埋藏深度为2 cm,以避免生物捕食造成的损失,种子发芽率可高达96%[30].在提高海草移植成功率方面,修复海草床植被的常用方法是草皮法、草块法和根状茎法.为了提高海草移植效率,澳大利亚研发了移植海草的专用机械设备ECOSUB1和ECOSUB2[31-32],提高了海草移植的机械化水平.另外,海草移植后容易被海浪冲走,因此还需对其进行固定.目前常用的海草移植固定方法有订书钉法、框架法和贝壳法.研究发现[33],采用订书钉法把海草两端固定于潮滩底部,4个月后海草的覆盖率可达到57.8%.被修复的海草床的固碳能力与天然海草床相近.例如:在澳大利亚西澳大利亚州牡蛎港的海岸移植海草18年后,海草床的碳埋藏率达到了0.3 Mg/(hm2·a)[34]. ...
Impact of seagrass loss and subsequent revegetation on carbon sequestration and stocks
2
2015
... 受到水产养殖、船舶运输和有害藻华的影响,海草大面积减少,导致裸露的沉积物中封存的有机碳加速分解.海草床植被修复型增汇技术主要通过提高海草种子萌芽率和海草移植成功率的方式来恢复海草床的碳封存能力.在提高海草种子萌芽率方面,已有多种因地制宜的技术被成功研发.例如:用淡水浸泡种子或在种子表面制造划痕,来有效诱导海草种子发芽[29];甲壳动物对海草种子的捕食会降低海草床的恢复成功率,因此将海菖蒲(Enhalus acoroides)种子装入网袋且使其埋藏深度为2 cm,以避免生物捕食造成的损失,种子发芽率可高达96%[30].在提高海草移植成功率方面,修复海草床植被的常用方法是草皮法、草块法和根状茎法.为了提高海草移植效率,澳大利亚研发了移植海草的专用机械设备ECOSUB1和ECOSUB2[31-32],提高了海草移植的机械化水平.另外,海草移植后容易被海浪冲走,因此还需对其进行固定.目前常用的海草移植固定方法有订书钉法、框架法和贝壳法.研究发现[33],采用订书钉法把海草两端固定于潮滩底部,4个月后海草的覆盖率可达到57.8%.被修复的海草床的固碳能力与天然海草床相近.例如:在澳大利亚西澳大利亚州牡蛎港的海岸移植海草18年后,海草床的碳埋藏率达到了0.3 Mg/(hm2·a)[34]. ...
... 移植海草(澳大利亚牡蛎港[34]) ...
Restoration of tropical seagrass beds using wild bird fertilization and sediment regrading
2
2018
... 船舶螺旋桨转动、船体搁浅及锚泊都会对浅海区海草床沉积物的结构和功能造成破坏,导致沉积物中的养分流失.海草床生境改善型增汇技术则通过施肥、缓解营养限制以及改善生长环境等方式,快速促进海草床植被的生长及其面积的增加,从而提高海草光合固碳量.例如:建立鸟桩(用于积累粪便的鸟类栖息桩)后,由于鸟类粪便中富含的磷等营养物质可以促进海草生长,2年后浅滩草(Halodule wrightii)的覆盖率接近于天然海草床[35].不过,鸟桩也可能会造成对海草床的过营养化,破坏群落演替并且增加附生植物生物量.与在海草床中引入更多鸟类相似,研究人员还研发了人工添加鸟粪石的海草床生境改善技术.该技术的原理是,经过废水处理后回收的鸟粪石不溶于水,而在接触海草根系分泌的有机酸后会快速释放磷.研究发现[36],添加鸟粪石后,海草床生物量增加了10%~60%,相比于常见的控释肥料,采用鸟粪石人工添加技术的生境改善效果更佳,且其释放的氮和磷更少. ...
... 通过鸟类栖息的木桩施加鸟粪肥料(美国佛罗里达群岛)[35] ...
Use of a wastewater recovery product (struvite) to enhance subtropical seagrass restoration
1
2022
... 船舶螺旋桨转动、船体搁浅及锚泊都会对浅海区海草床沉积物的结构和功能造成破坏,导致沉积物中的养分流失.海草床生境改善型增汇技术则通过施肥、缓解营养限制以及改善生长环境等方式,快速促进海草床植被的生长及其面积的增加,从而提高海草光合固碳量.例如:建立鸟桩(用于积累粪便的鸟类栖息桩)后,由于鸟类粪便中富含的磷等营养物质可以促进海草生长,2年后浅滩草(Halodule wrightii)的覆盖率接近于天然海草床[35].不过,鸟桩也可能会造成对海草床的过营养化,破坏群落演替并且增加附生植物生物量.与在海草床中引入更多鸟类相似,研究人员还研发了人工添加鸟粪石的海草床生境改善技术.该技术的原理是,经过废水处理后回收的鸟粪石不溶于水,而在接触海草根系分泌的有机酸后会快速释放磷.研究发现[36],添加鸟粪石后,海草床生物量增加了10%~60%,相比于常见的控释肥料,采用鸟粪石人工添加技术的生境改善效果更佳,且其释放的氮和磷更少. ...
Seaweed blue carbon: Ready? or not?
2
2023
... 近岸海藻场增汇技术即通过优选藻种和改良养殖技术来缓解海藻场固碳的部分限制作用.例如:选择养殖产量较高的海藻如海带(Laminaria),其每年的碳固定量超过30.0 Mg/hm2[37].2010—2020年,全球的海藻养殖产量约增加了1.5×107 Mg,海藻养殖密度增大,导致海藻营养缺乏,生长受阻.可采用气幕式人工上升流装备将海底富营养盐的海水输送到海面,辅助浮球下的海带生长.目前,已在山东青岛的鳌山湾海域开展国内首个人工上升流增汇示范工程,通过提升上升流的涌升效率,每株海带平均增产36.1 g,CO2吸收量约增加了1.6 Mg/hm2[38].日本制铁公司开发了一种由钢铁渣与废木材发酵形成的腐殖土混合而成的肥料,该肥料在2004年10月施用于北海道试点区域,8个月后发现试验区的海带生物量比非实验区增加了100多倍[39].光照限制是海藻养殖的严重瓶颈,特别在浑浊水域,大量的悬浮物会使水下辐照度降低,影响海藻的生长.利用波浪摩擦发电而为海藻提供光照是促进海藻生长和提高其碳汇效能的一种可行方案[40].实验室研究表明,利用功率为90 mW左右的波浪发电机补光装备为海带补光,可使海带的平均净生物量积累率和线性生长率显著提高,分别提高10.83±0.16%和9.12±0.68%[40]. ...
... 开发海藻养殖区域同样至关重要.海藻常用养殖装备为浮筏,其适用于风浪较小的近岸浅水区,在深水区浮筏的养殖锚绳会在恶劣海况下断裂.目前,离岸系泊式海藻牧场的构建备受关注,其突破了海藻在40 m水深内养殖的限制,扩大了海藻的养殖区域[41].在韩国南海郡,研究人员利用中水层绳索培养技术种植多年生褐藻昆布(Ecklonia cava),将海藻苗绳悬挂在水中,使海藻在水中生长.该技术的优点是可以避免海藻苗被海洋生物捕食,同时种植设备可以与其他装备(如海上风电场和平台)结合使用.该技术的试点农场每年约可以吸收10.0 Mg/hm2的CO2,有效减少了温室气体的排放[42].此外,海藻沉降被认为是可以实现永久碳封存的有效技术手段,如果可以将海藻下沉到海洋深处的无氧区域(水深>1 000 m),那么海藻有可能被长时间保存[37].因此,开发具有环境感知能力及主动沉降能力的智能化深远海海藻养殖装备是发展海藻碳汇的可行路径. ...
促进蓝碳增汇 给海洋牧场装上巨型“气泵”
2
... 近岸海藻场增汇技术即通过优选藻种和改良养殖技术来缓解海藻场固碳的部分限制作用.例如:选择养殖产量较高的海藻如海带(Laminaria),其每年的碳固定量超过30.0 Mg/hm2[37].2010—2020年,全球的海藻养殖产量约增加了1.5×107 Mg,海藻养殖密度增大,导致海藻营养缺乏,生长受阻.可采用气幕式人工上升流装备将海底富营养盐的海水输送到海面,辅助浮球下的海带生长.目前,已在山东青岛的鳌山湾海域开展国内首个人工上升流增汇示范工程,通过提升上升流的涌升效率,每株海带平均增产36.1 g,CO2吸收量约增加了1.6 Mg/hm2[38].日本制铁公司开发了一种由钢铁渣与废木材发酵形成的腐殖土混合而成的肥料,该肥料在2004年10月施用于北海道试点区域,8个月后发现试验区的海带生物量比非实验区增加了100多倍[39].光照限制是海藻养殖的严重瓶颈,特别在浑浊水域,大量的悬浮物会使水下辐照度降低,影响海藻的生长.利用波浪摩擦发电而为海藻提供光照是促进海藻生长和提高其碳汇效能的一种可行方案[40].实验室研究表明,利用功率为90 mW左右的波浪发电机补光装备为海带补光,可使海带的平均净生物量积累率和线性生长率显著提高,分别提高10.83±0.16%和9.12±0.68%[40]. ...
... (山东青岛鳌山湾)[38] ...
促进蓝碳增汇 给海洋牧场装上巨型“气泵”
2
... 近岸海藻场增汇技术即通过优选藻种和改良养殖技术来缓解海藻场固碳的部分限制作用.例如:选择养殖产量较高的海藻如海带(Laminaria),其每年的碳固定量超过30.0 Mg/hm2[37].2010—2020年,全球的海藻养殖产量约增加了1.5×107 Mg,海藻养殖密度增大,导致海藻营养缺乏,生长受阻.可采用气幕式人工上升流装备将海底富营养盐的海水输送到海面,辅助浮球下的海带生长.目前,已在山东青岛的鳌山湾海域开展国内首个人工上升流增汇示范工程,通过提升上升流的涌升效率,每株海带平均增产36.1 g,CO2吸收量约增加了1.6 Mg/hm2[38].日本制铁公司开发了一种由钢铁渣与废木材发酵形成的腐殖土混合而成的肥料,该肥料在2004年10月施用于北海道试点区域,8个月后发现试验区的海带生物量比非实验区增加了100多倍[39].光照限制是海藻养殖的严重瓶颈,特别在浑浊水域,大量的悬浮物会使水下辐照度降低,影响海藻的生长.利用波浪摩擦发电而为海藻提供光照是促进海藻生长和提高其碳汇效能的一种可行方案[40].实验室研究表明,利用功率为90 mW左右的波浪发电机补光装备为海带补光,可使海带的平均净生物量积累率和线性生长率显著提高,分别提高10.83±0.16%和9.12±0.68%[40]. ...
... (山东青岛鳌山湾)[38] ...
The recycling and reuse of steelmaking slags: A review
1
2019
... 近岸海藻场增汇技术即通过优选藻种和改良养殖技术来缓解海藻场固碳的部分限制作用.例如:选择养殖产量较高的海藻如海带(Laminaria),其每年的碳固定量超过30.0 Mg/hm2[37].2010—2020年,全球的海藻养殖产量约增加了1.5×107 Mg,海藻养殖密度增大,导致海藻营养缺乏,生长受阻.可采用气幕式人工上升流装备将海底富营养盐的海水输送到海面,辅助浮球下的海带生长.目前,已在山东青岛的鳌山湾海域开展国内首个人工上升流增汇示范工程,通过提升上升流的涌升效率,每株海带平均增产36.1 g,CO2吸收量约增加了1.6 Mg/hm2[38].日本制铁公司开发了一种由钢铁渣与废木材发酵形成的腐殖土混合而成的肥料,该肥料在2004年10月施用于北海道试点区域,8个月后发现试验区的海带生物量比非实验区增加了100多倍[39].光照限制是海藻养殖的严重瓶颈,特别在浑浊水域,大量的悬浮物会使水下辐照度降低,影响海藻的生长.利用波浪摩擦发电而为海藻提供光照是促进海藻生长和提高其碳汇效能的一种可行方案[40].实验室研究表明,利用功率为90 mW左右的波浪发电机补光装备为海带补光,可使海带的平均净生物量积累率和线性生长率显著提高,分别提高10.83±0.16%和9.12±0.68%[40]. ...
Self-powered and self-sensing blue carbon ecosystems by hybrid fur triboelectric nanogenerators(F-TENG)
2
2024
... 近岸海藻场增汇技术即通过优选藻种和改良养殖技术来缓解海藻场固碳的部分限制作用.例如:选择养殖产量较高的海藻如海带(Laminaria),其每年的碳固定量超过30.0 Mg/hm2[37].2010—2020年,全球的海藻养殖产量约增加了1.5×107 Mg,海藻养殖密度增大,导致海藻营养缺乏,生长受阻.可采用气幕式人工上升流装备将海底富营养盐的海水输送到海面,辅助浮球下的海带生长.目前,已在山东青岛的鳌山湾海域开展国内首个人工上升流增汇示范工程,通过提升上升流的涌升效率,每株海带平均增产36.1 g,CO2吸收量约增加了1.6 Mg/hm2[38].日本制铁公司开发了一种由钢铁渣与废木材发酵形成的腐殖土混合而成的肥料,该肥料在2004年10月施用于北海道试点区域,8个月后发现试验区的海带生物量比非实验区增加了100多倍[39].光照限制是海藻养殖的严重瓶颈,特别在浑浊水域,大量的悬浮物会使水下辐照度降低,影响海藻的生长.利用波浪摩擦发电而为海藻提供光照是促进海藻生长和提高其碳汇效能的一种可行方案[40].实验室研究表明,利用功率为90 mW左右的波浪发电机补光装备为海带补光,可使海带的平均净生物量积累率和线性生长率显著提高,分别提高10.83±0.16%和9.12±0.68%[40]. ...
... [40]. ...
离岸系泊式海藻牧场的动力响应分析
1
2023
... 开发海藻养殖区域同样至关重要.海藻常用养殖装备为浮筏,其适用于风浪较小的近岸浅水区,在深水区浮筏的养殖锚绳会在恶劣海况下断裂.目前,离岸系泊式海藻牧场的构建备受关注,其突破了海藻在40 m水深内养殖的限制,扩大了海藻的养殖区域[41].在韩国南海郡,研究人员利用中水层绳索培养技术种植多年生褐藻昆布(Ecklonia cava),将海藻苗绳悬挂在水中,使海藻在水中生长.该技术的优点是可以避免海藻苗被海洋生物捕食,同时种植设备可以与其他装备(如海上风电场和平台)结合使用.该技术的试点农场每年约可以吸收10.0 Mg/hm2的CO2,有效减少了温室气体的排放[42].此外,海藻沉降被认为是可以实现永久碳封存的有效技术手段,如果可以将海藻下沉到海洋深处的无氧区域(水深>1 000 m),那么海藻有可能被长时间保存[37].因此,开发具有环境感知能力及主动沉降能力的智能化深远海海藻养殖装备是发展海藻碳汇的可行路径. ...
离岸系泊式海藻牧场的动力响应分析
1
2023
... 开发海藻养殖区域同样至关重要.海藻常用养殖装备为浮筏,其适用于风浪较小的近岸浅水区,在深水区浮筏的养殖锚绳会在恶劣海况下断裂.目前,离岸系泊式海藻牧场的构建备受关注,其突破了海藻在40 m水深内养殖的限制,扩大了海藻的养殖区域[41].在韩国南海郡,研究人员利用中水层绳索培养技术种植多年生褐藻昆布(Ecklonia cava),将海藻苗绳悬挂在水中,使海藻在水中生长.该技术的优点是可以避免海藻苗被海洋生物捕食,同时种植设备可以与其他装备(如海上风电场和平台)结合使用.该技术的试点农场每年约可以吸收10.0 Mg/hm2的CO2,有效减少了温室气体的排放[42].此外,海藻沉降被认为是可以实现永久碳封存的有效技术手段,如果可以将海藻下沉到海洋深处的无氧区域(水深>1 000 m),那么海藻有可能被长时间保存[37].因此,开发具有环境感知能力及主动沉降能力的智能化深远海海藻养殖装备是发展海藻碳汇的可行路径. ...
Installing kelp forests/seaweed beds for mitigation and adaptation against global warming: Korean project overview
2
2013
... 开发海藻养殖区域同样至关重要.海藻常用养殖装备为浮筏,其适用于风浪较小的近岸浅水区,在深水区浮筏的养殖锚绳会在恶劣海况下断裂.目前,离岸系泊式海藻牧场的构建备受关注,其突破了海藻在40 m水深内养殖的限制,扩大了海藻的养殖区域[41].在韩国南海郡,研究人员利用中水层绳索培养技术种植多年生褐藻昆布(Ecklonia cava),将海藻苗绳悬挂在水中,使海藻在水中生长.该技术的优点是可以避免海藻苗被海洋生物捕食,同时种植设备可以与其他装备(如海上风电场和平台)结合使用.该技术的试点农场每年约可以吸收10.0 Mg/hm2的CO2,有效减少了温室气体的排放[42].此外,海藻沉降被认为是可以实现永久碳封存的有效技术手段,如果可以将海藻下沉到海洋深处的无氧区域(水深>1 000 m),那么海藻有可能被长时间保存[37].因此,开发具有环境感知能力及主动沉降能力的智能化深远海海藻养殖装备是发展海藻碳汇的可行路径. ...
... 南道南海郡)[42] ...
滨海电厂温排水温升对红树林分布扩散的影响
1
2024
... 核电站的换热装置通过循环加热冷却水的方式进行热量交换,并将加热后的冷却水排入邻近海域,致使周围海水的温度升高.该受热的海水称为温排水.红树植物多具有嗜热性,其分布受温度因素 (包括气温、水温)的主导影响,因此,利用温排水帮助红树林御寒,已成为一种具有多重效益的碳汇手段.针对广西防城港核电站温排水区域红树林的研究表明[43],受温排水影响的红树林在各个温升区均有不同程度的自然生长扩散现象.但是,由于水温升高,温排水的溶解氧含量较低,同时水体中残留的人为添加的除生剂(如氯气)对红树林的生长扩散具有一定的影响,因此需要有智能化的温排水监测、处理及排放装备作保障. ...
滨海电厂温排水温升对红树林分布扩散的影响
1
2024
... 核电站的换热装置通过循环加热冷却水的方式进行热量交换,并将加热后的冷却水排入邻近海域,致使周围海水的温度升高.该受热的海水称为温排水.红树植物多具有嗜热性,其分布受温度因素 (包括气温、水温)的主导影响,因此,利用温排水帮助红树林御寒,已成为一种具有多重效益的碳汇手段.针对广西防城港核电站温排水区域红树林的研究表明[43],受温排水影响的红树林在各个温升区均有不同程度的自然生长扩散现象.但是,由于水温升高,温排水的溶解氧含量较低,同时水体中残留的人为添加的除生剂(如氯气)对红树林的生长扩散具有一定的影响,因此需要有智能化的温排水监测、处理及排放装备作保障. ...
Aquaculture Perspective of Multi-Use Sites in the Open Ocean: The Untapped Potential for Marine Resources in the Anthropocene
2
2017
... 海藻场与风电场的结合可以使两者共享运维.研究表明[44],海水养殖与风电场相结合,可以使风电场的运维成本降低10%.风机基座可用于海藻养殖装置的附着固定,风电场为海藻场提供无航区域,使海藻养殖由近岸向开放海域延伸.将海藻场与近海风电场结合的计划早在2000年在德国北海提出.考虑到不同于近岸的复杂海况,风电场内海藻养殖装备由长线结构和环形结构发展为可自主下潜的笼状结构,其对风浪和复杂海况的抵御效果逐步得到提升[44].在中国,风电场远离海岸,为了使海藻养殖产业与风电产业相结合而产生显著的耦合效果,技术装备需要进一步提升. ...
... [44].在中国,风电场远离海岸,为了使海藻养殖产业与风电产业相结合而产生显著的耦合效果,技术装备需要进一步提升. ...
Expert review of the science underlying nature-based climate solutions
2
2024
... 滨海蓝碳增汇的技术、装备和效益如表1所示.由表可知:红树林、盐沼、海草床、海藻场的CO2吸收率分别为3.0×108~3.6×108,5.0×107~7.0×107,1.3×108~1.6×108,3.0×108 Mg/a[45];红树林增汇项目的成本约为23.0~3.7×105美元/(hm2·a),主要包括植物及其种植、施工、维护、运输等成本,其具有可观的投资回报,后续的经济效益约为146.0~5.1×105美元/(hm2·a)[46];盐沼增汇项目的成本约为1.2×103~3.3×105 美元/(hm2·a),后期的经济效益为1.0×103~3.1×105 美元/(hm2·a)[47];海草床增汇项目的成本为1.2×104~7.0×105 美元/(hm2·a),后期的经济效益为1.9×103~3.6×105 美元/(hm2·a)[48];海藻场增汇项目的成本为3.4×104~2.2×105美元/(hm2·a),后期的经济效益为6.4×104~1.4×105美元/(hm2·a)[49].滨海蓝碳增汇项目大约需要投入3~10年,前期的经济净收益较少,后期项目完成后每年都有较高且稳定的经济效益. ...
... (Mg/a)[45] ...
A meta-analysis of the ecological and economic outcomes of mangrove restoration
3
2021
... 滨海蓝碳增汇的技术、装备和效益如表1所示.由表可知:红树林、盐沼、海草床、海藻场的CO2吸收率分别为3.0×108~3.6×108,5.0×107~7.0×107,1.3×108~1.6×108,3.0×108 Mg/a[45];红树林增汇项目的成本约为23.0~3.7×105美元/(hm2·a),主要包括植物及其种植、施工、维护、运输等成本,其具有可观的投资回报,后续的经济效益约为146.0~5.1×105美元/(hm2·a)[46];盐沼增汇项目的成本约为1.2×103~3.3×105 美元/(hm2·a),后期的经济效益为1.0×103~3.1×105 美元/(hm2·a)[47];海草床增汇项目的成本为1.2×104~7.0×105 美元/(hm2·a),后期的经济效益为1.9×103~3.6×105 美元/(hm2·a)[48];海藻场增汇项目的成本为3.4×104~2.2×105美元/(hm2·a),后期的经济效益为6.4×104~1.4×105美元/(hm2·a)[49].滨海蓝碳增汇项目大约需要投入3~10年,前期的经济净收益较少,后期项目完成后每年都有较高且稳定的经济效益. ...
... (hm2·a)][46-49] ...
... (hm2·a)][46-49] ...
Public perceptions of blue carbon in South Korea: Findings from a choice experiment
1
2022
... 滨海蓝碳增汇的技术、装备和效益如表1所示.由表可知:红树林、盐沼、海草床、海藻场的CO2吸收率分别为3.0×108~3.6×108,5.0×107~7.0×107,1.3×108~1.6×108,3.0×108 Mg/a[45];红树林增汇项目的成本约为23.0~3.7×105美元/(hm2·a),主要包括植物及其种植、施工、维护、运输等成本,其具有可观的投资回报,后续的经济效益约为146.0~5.1×105美元/(hm2·a)[46];盐沼增汇项目的成本约为1.2×103~3.3×105 美元/(hm2·a),后期的经济效益为1.0×103~3.1×105 美元/(hm2·a)[47];海草床增汇项目的成本为1.2×104~7.0×105 美元/(hm2·a),后期的经济效益为1.9×103~3.6×105 美元/(hm2·a)[48];海藻场增汇项目的成本为3.4×104~2.2×105美元/(hm2·a),后期的经济效益为6.4×104~1.4×105美元/(hm2·a)[49].滨海蓝碳增汇项目大约需要投入3~10年,前期的经济净收益较少,后期项目完成后每年都有较高且稳定的经济效益. ...
Assessing the CO2 capture potential of seagrass restoration projects
1
2013
... 滨海蓝碳增汇的技术、装备和效益如表1所示.由表可知:红树林、盐沼、海草床、海藻场的CO2吸收率分别为3.0×108~3.6×108,5.0×107~7.0×107,1.3×108~1.6×108,3.0×108 Mg/a[45];红树林增汇项目的成本约为23.0~3.7×105美元/(hm2·a),主要包括植物及其种植、施工、维护、运输等成本,其具有可观的投资回报,后续的经济效益约为146.0~5.1×105美元/(hm2·a)[46];盐沼增汇项目的成本约为1.2×103~3.3×105 美元/(hm2·a),后期的经济效益为1.0×103~3.1×105 美元/(hm2·a)[47];海草床增汇项目的成本为1.2×104~7.0×105 美元/(hm2·a),后期的经济效益为1.9×103~3.6×105 美元/(hm2·a)[48];海藻场增汇项目的成本为3.4×104~2.2×105美元/(hm2·a),后期的经济效益为6.4×104~1.4×105美元/(hm2·a)[49].滨海蓝碳增汇项目大约需要投入3~10年,前期的经济净收益较少,后期项目完成后每年都有较高且稳定的经济效益. ...
The value of ecosystem services in global marine kelp forests
3
2023
... 滨海蓝碳增汇的技术、装备和效益如表1所示.由表可知:红树林、盐沼、海草床、海藻场的CO2吸收率分别为3.0×108~3.6×108,5.0×107~7.0×107,1.3×108~1.6×108,3.0×108 Mg/a[45];红树林增汇项目的成本约为23.0~3.7×105美元/(hm2·a),主要包括植物及其种植、施工、维护、运输等成本,其具有可观的投资回报,后续的经济效益约为146.0~5.1×105美元/(hm2·a)[46];盐沼增汇项目的成本约为1.2×103~3.3×105 美元/(hm2·a),后期的经济效益为1.0×103~3.1×105 美元/(hm2·a)[47];海草床增汇项目的成本为1.2×104~7.0×105 美元/(hm2·a),后期的经济效益为1.9×103~3.6×105 美元/(hm2·a)[48];海藻场增汇项目的成本为3.4×104~2.2×105美元/(hm2·a),后期的经济效益为6.4×104~1.4×105美元/(hm2·a)[49].滨海蓝碳增汇项目大约需要投入3~10年,前期的经济净收益较少,后期项目完成后每年都有较高且稳定的经济效益. ...
... (hm2·a)][46-49] ...
... (hm2·a)][46-49] ...
Integrated marsh management (IMM): A new perspective on mosquito control and best management practices for salt marsh restoration
1
2012
... 重新造林后,红树林湿地的螃蟹、多毛类蠕虫和两栖动物等生物群落的密度增加,此外,红树林水文连通性恢复后,为以潮间带底栖动物为食的迁徙滨鸟和濒危鱼类提供了适宜的生存环境.美国纽约长岛的退化盐沼修复后,盐沼本土植被覆盖率从48%增加到78%,加拿大底鳉(Fundulus heteroclitus)、羊头鲷(Cyprinodon variegatus)、银鳃蟹(Menidia spp.)和蓝蟹(Callinectes sapidus)的丰度显著增加,滨鸟、水禽、雀鸟和猛禽的数量分别增加了110%、62%、84%和220%[50].美国得克萨斯州海岸移植海草10年后,鸟、无脊椎动物、双壳类动物和鱼的种类显著增加[51].在加勒比海的海藻养殖区域发现鱼类和无脊椎动物的种类分别增加了25倍和7倍,其中杜父鱼 (Anisotremus sp.)和沙丁鱼(Harengula sp.)的数量分别增加了4倍和2倍,并且以海藻为食的蝾螺(Turbinidae)和绿宝石蟹(Mithraculus sculptus)等动物大量存在,与周围海域相比,海藻养殖区域的生物多样性更显著[52]. ...
Longevity and sustainability of tropical and subtropical restored seagrass beds among Atlantic, Pacific, and Indian Oceans
1
2020
... 重新造林后,红树林湿地的螃蟹、多毛类蠕虫和两栖动物等生物群落的密度增加,此外,红树林水文连通性恢复后,为以潮间带底栖动物为食的迁徙滨鸟和濒危鱼类提供了适宜的生存环境.美国纽约长岛的退化盐沼修复后,盐沼本土植被覆盖率从48%增加到78%,加拿大底鳉(Fundulus heteroclitus)、羊头鲷(Cyprinodon variegatus)、银鳃蟹(Menidia spp.)和蓝蟹(Callinectes sapidus)的丰度显著增加,滨鸟、水禽、雀鸟和猛禽的数量分别增加了110%、62%、84%和220%[50].美国得克萨斯州海岸移植海草10年后,鸟、无脊椎动物、双壳类动物和鱼的种类显著增加[51].在加勒比海的海藻养殖区域发现鱼类和无脊椎动物的种类分别增加了25倍和7倍,其中杜父鱼 (Anisotremus sp.)和沙丁鱼(Harengula sp.)的数量分别增加了4倍和2倍,并且以海藻为食的蝾螺(Turbinidae)和绿宝石蟹(Mithraculus sculptus)等动物大量存在,与周围海域相比,海藻养殖区域的生物多样性更显著[52]. ...
Tropical seaweeds for human food, their cultivation and its effect on biodiversity enrichment
1
2015
... 重新造林后,红树林湿地的螃蟹、多毛类蠕虫和两栖动物等生物群落的密度增加,此外,红树林水文连通性恢复后,为以潮间带底栖动物为食的迁徙滨鸟和濒危鱼类提供了适宜的生存环境.美国纽约长岛的退化盐沼修复后,盐沼本土植被覆盖率从48%增加到78%,加拿大底鳉(Fundulus heteroclitus)、羊头鲷(Cyprinodon variegatus)、银鳃蟹(Menidia spp.)和蓝蟹(Callinectes sapidus)的丰度显著增加,滨鸟、水禽、雀鸟和猛禽的数量分别增加了110%、62%、84%和220%[50].美国得克萨斯州海岸移植海草10年后,鸟、无脊椎动物、双壳类动物和鱼的种类显著增加[51].在加勒比海的海藻养殖区域发现鱼类和无脊椎动物的种类分别增加了25倍和7倍,其中杜父鱼 (Anisotremus sp.)和沙丁鱼(Harengula sp.)的数量分别增加了4倍和2倍,并且以海藻为食的蝾螺(Turbinidae)和绿宝石蟹(Mithraculus sculptus)等动物大量存在,与周围海域相比,海藻养殖区域的生物多样性更显著[52]. ...






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}